

A Challenging Pie to Splice: Drugging the Spliceosome
- PMID: 28371109
- PMCID: PMC6311392
- DOI: 10.1002/anie.201701065
A Challenging Pie to Splice: Drugging the Spliceosome
Abstract
Since its discovery in 1977, the study of alternative RNA splicing has revealed a plethora of mechanisms that had never before been documented in nature. Understanding these transitions and their outcome at the level of the cell and organism has become one of the great frontiers of modern chemical biology. Until 2007, this field remained in the hands of RNA biologists. However, the recent identification of natural product and synthetic modulators of RNA splicing has opened new access to this field, allowing for the first time a chemical-based interrogation of RNA splicing processes. Simultaneously, we have begun to understand the vital importance of splicing in disease, which offers a new platform for molecular discovery and therapy. As with many natural systems, gaining clear mechanistic detail at the molecular level is key towards understanding the operation of any biological machine. This minireview presents recent lessons learned in this emerging field of RNA splicing chemistry and chemical biology.
Keywords: RNA splicing; chemical biology; drug discovery; inhibitors; spliceosome.
© 2017 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim.
Conflict of interest statement
Conflict of interest
The authors declare no conflict of interest.
Figures

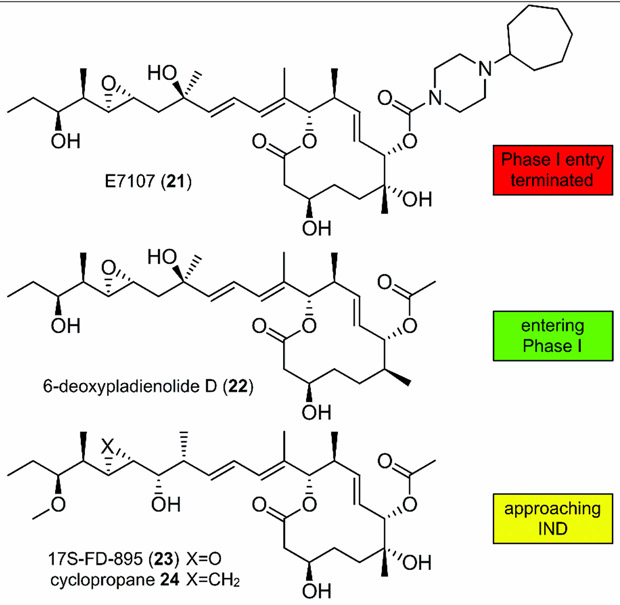
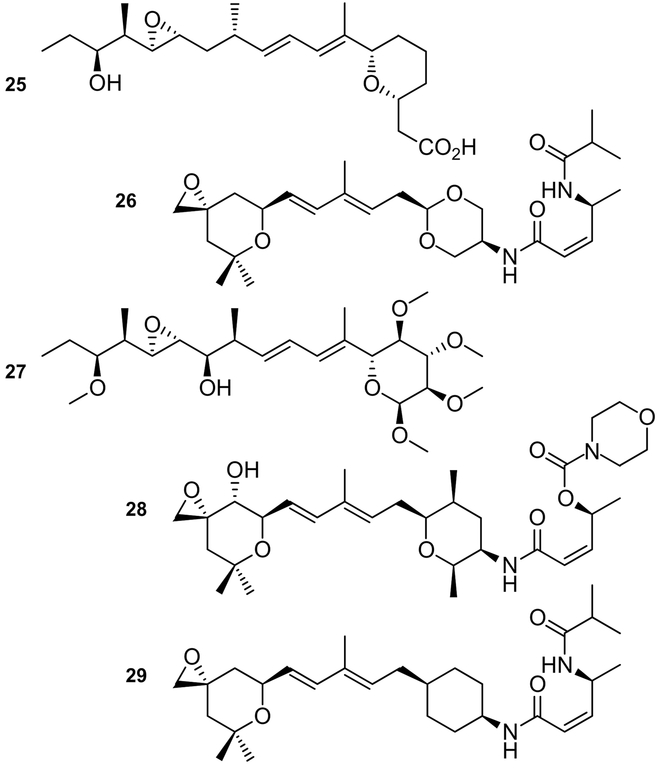
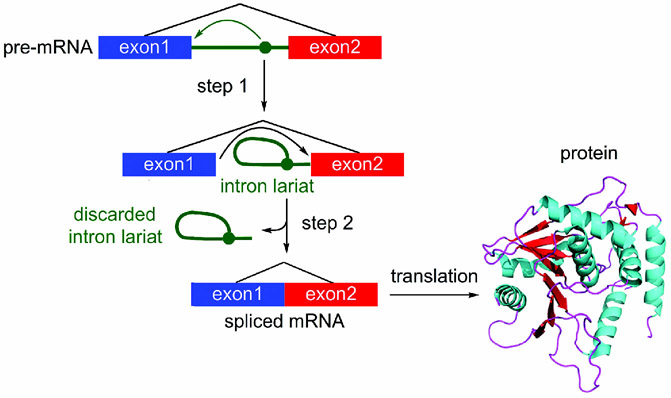
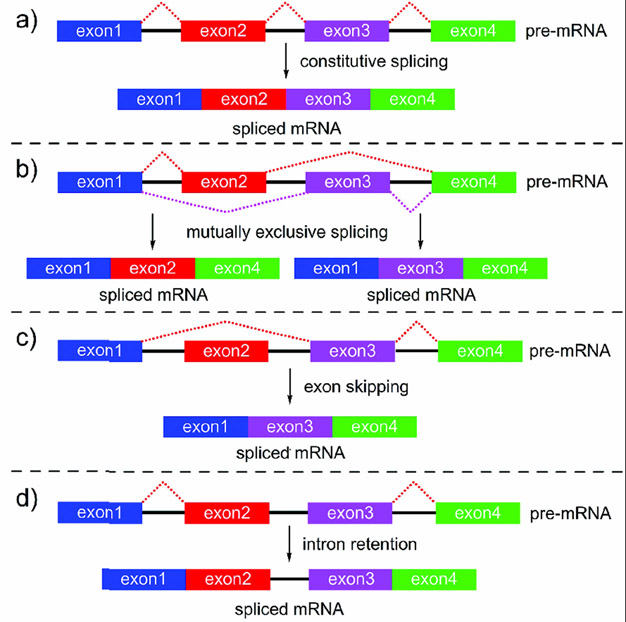
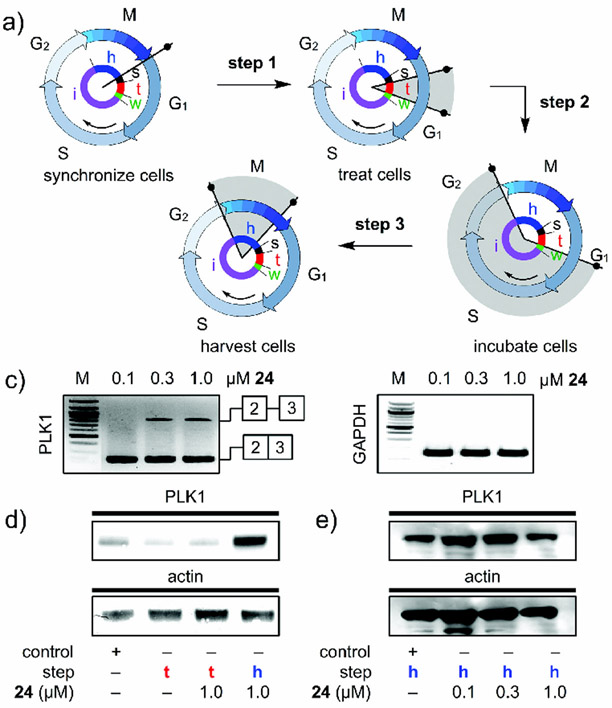
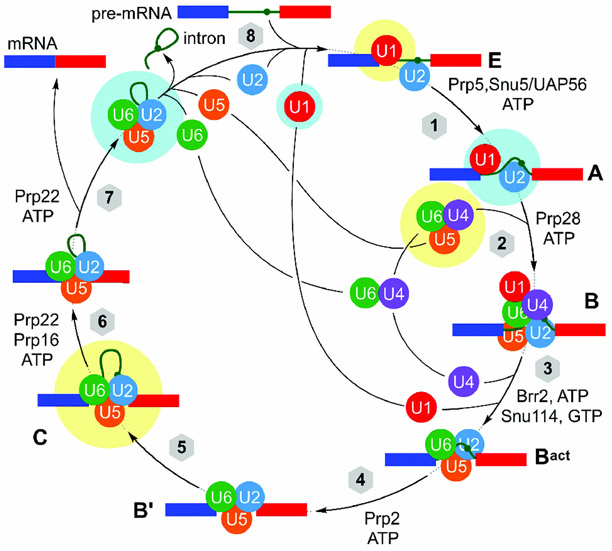
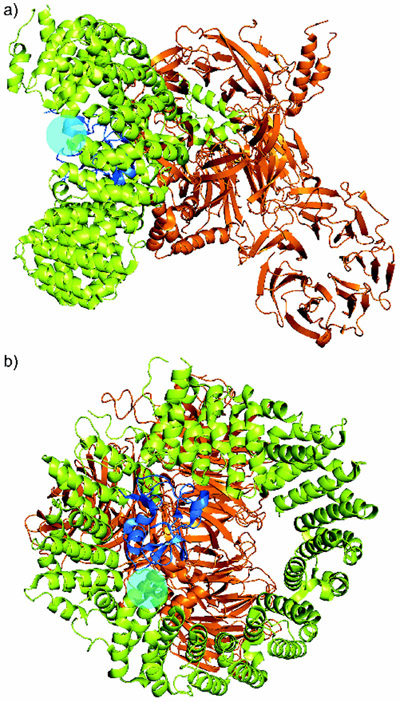
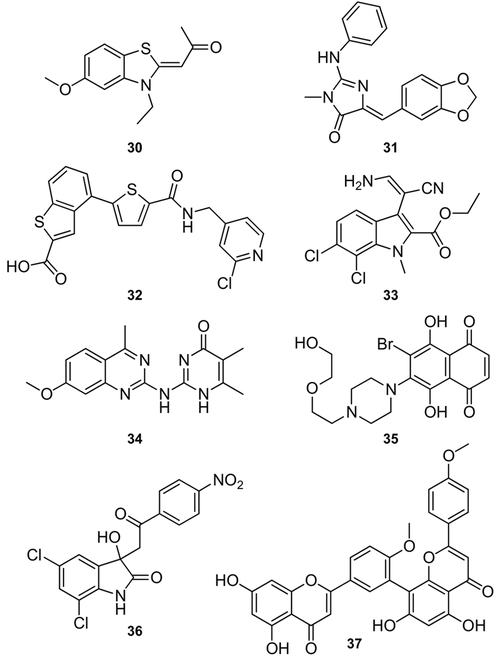
References
-
- Kaida D.,Motoyoshi H, Tashiro E, Nojima T, Hagiwara M, Ishigami K, Watanabe H, Kitahara T, Yoshida T, Nakajima H, Tani T, Horinouchi S, Yoshida M, Nat. Chem. Biol. 2007, 3, 576.; - PubMed
- Kotake Y., Sagane K, Owa T, Mimori-Kiyosue Y, Shimizu H, Uesugi M, Ishihama Y, Iwata M, Mizui Y, Nat. Chem. Biol. 2007, 3, 570. - PubMed
-
- Isaac BG., Ayer SW, Elliott RC, Stonard RJ, J. Org. Chem. 1992, 57, 7220.;
- Sakai Y, Yoshida T, Ochiai K, Uosaki Y, Saitoh Y, Tanaka F, Akiyama T, Akinaga S, Mizukami T, J. Antibiot. 2002, 55, 855. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
