

Phosphatidylinositol 3-phosphate-binding protein AtPH1 controls the localization of the metal transporter NRAMP1 in Arabidopsis
- PMID: 28373552
- PMCID: PMC5402440
- DOI: 10.1073/pnas.1702975114
Phosphatidylinositol 3-phosphate-binding protein AtPH1 controls the localization of the metal transporter NRAMP1 in Arabidopsis
Abstract
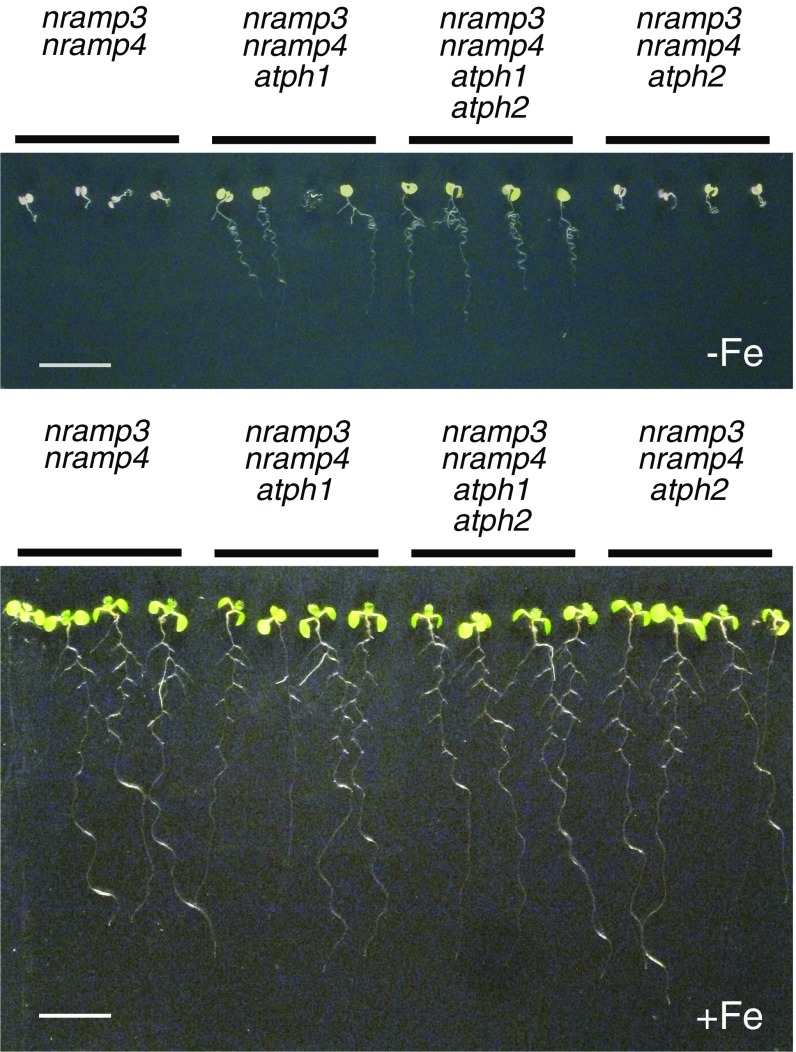
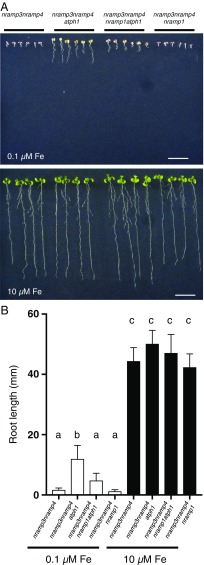
"Too much of a good thing" perfectly describes the dilemma that living organisms face with metals. The tight control of metal homeostasis in cells depends on the trafficking of metal transporters between membranes of different compartments. However, the mechanisms regulating the location of transport proteins are still largely unknown. Developing Arabidopsis thaliana seedlings require the natural resistance-associated macrophage proteins (NRAMP3 and NRAMP4) transporters to remobilize iron from seed vacuolar stores and thereby acquire photosynthetic competence. Here, we report that mutations in the pleckstrin homology (PH) domain-containing protein AtPH1 rescue the iron-deficient phenotype of nramp3nramp4 Our results indicate that AtPH1 binds phosphatidylinositol 3-phosphate (PI3P) in vivo and acts in the late endosome compartment. We further show that loss of AtPH1 function leads to the mislocalization of the metal uptake transporter NRAMP1 to the vacuole, providing a rationale for the reversion of nramp3nramp4 phenotypes. This work identifies a PH domain protein as a regulator of plant metal transporter localization, providing evidence that PH domain proteins may be effectors of PI3P for protein sorting.
Keywords: NRAMP; late endosome; metal transport; phosphatidylinositol 3-phosphate; vacuole.
Conflict of interest statement
The authors declare no conflict of interest.
Figures















References
-
- Briat JF, Dubos C, Gaymard F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 2015;20:33–40. - PubMed
-
- Murgia I, Arosio P, Tarantino D, Soave C. Biofortification for combating ‘hidden hunger’ for iron. Trends Plant Sci. 2012;17:47–55. - PubMed
-
- Robinson NJ, Procter CM, Connolly EL, Guerinot ML. A ferric-chelate reductase for iron uptake from soils. Nature. 1999;397:694–697. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
