

A model for the irradiance responses of photosynthesis
- PMID: 28374429
- PMCID: PMC5575564
- DOI: 10.1111/ppl.12572
A model for the irradiance responses of photosynthesis
Abstract
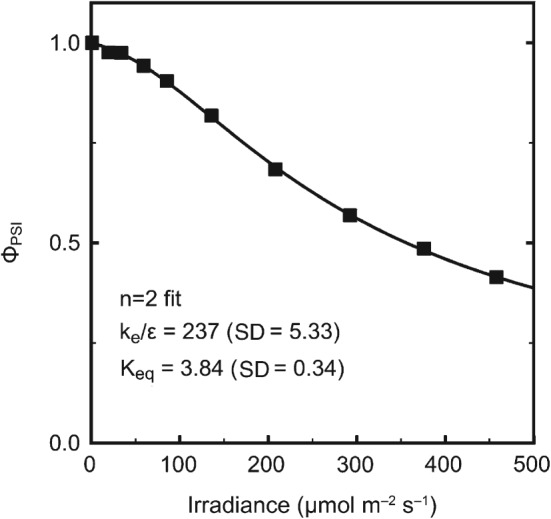
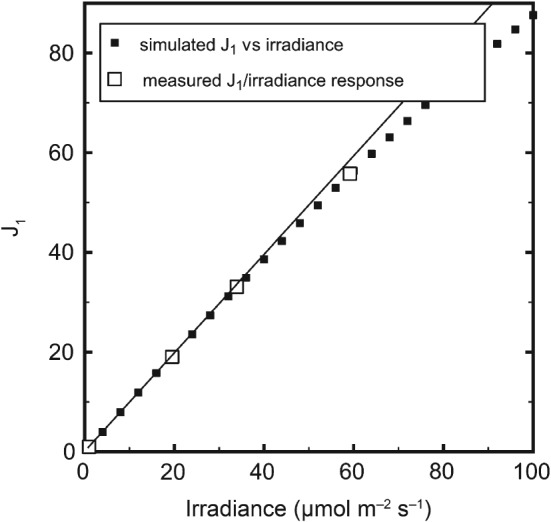
The analysis of the irradiance responses of photosynthetic processes, such as the quantum efficiencies of electron transport by photosystems I and II (PSI and PSII) or the rate of carbon dioxide fixation, is limited by the lack of mechanistically based analytical model for these processes. Starting with a model of P700 redox state, we develop a series of analytical functions which can be used to fit the irradiance responses of the quantum yields for electron transport by PSI and PSII, the irradiance responses of electron transport by PSI and PSII, and even the irradiance response of the fixation rate of carbon dioxide. These functions depend on two or three parameters so they can be fit to typical irradiance response data. We illustrate by example the use of these functions in various applications and discuss further use and development of the basic model described in detail here.
© 2017 The Authors. Physiologia Plantarum published by John Wiley & Sons Ltd on behalf of Scandinavian Plant Physiology Society.
Figures















References
-
- Anderson JM (1992) Cytochrome b6f complex: dynamic molecular organization, function and acclimation. Photosynth Res 34: 341–357 - PubMed
-
- Badeck FW (1995) Intra‐leaf gradient of assimilation rate and optimal allocation of canopy nitrogen: a model on the implications of the use of homogeneous assimilation functions. Funct Plant Biol 22: 425–439
-
- Baker NR, Harbinson J, Kramer DM (2007) Determining the limitations and regulation of photosynthetic energy transduction in leaves. Plant Cell Environ 30: 1107–1125 - PubMed
-
- Björkman O, Demmig B (1987) Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 170: 489–504 - PubMed
-
- Cape JL, Bowman MK, Kramer DM (2006) Understanding the cytochrome bc complexes by what they don't do. The Q‐cycle at 30. Trends Plant Sci 11: 46–55 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
