

Bovine and murine models highlight novel roles for SLC25A46 in mitochondrial dynamics and metabolism, with implications for human and animal health
- PMID: 28376083
- PMCID: PMC5380314
- DOI: 10.1371/journal.pgen.1006597
Bovine and murine models highlight novel roles for SLC25A46 in mitochondrial dynamics and metabolism, with implications for human and animal health
Abstract
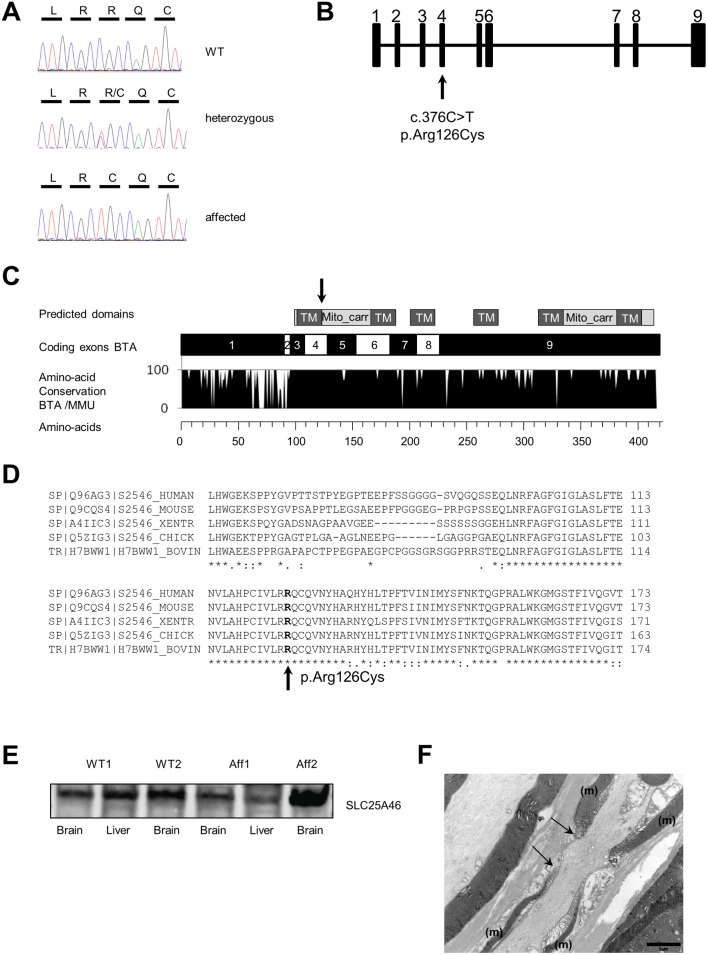
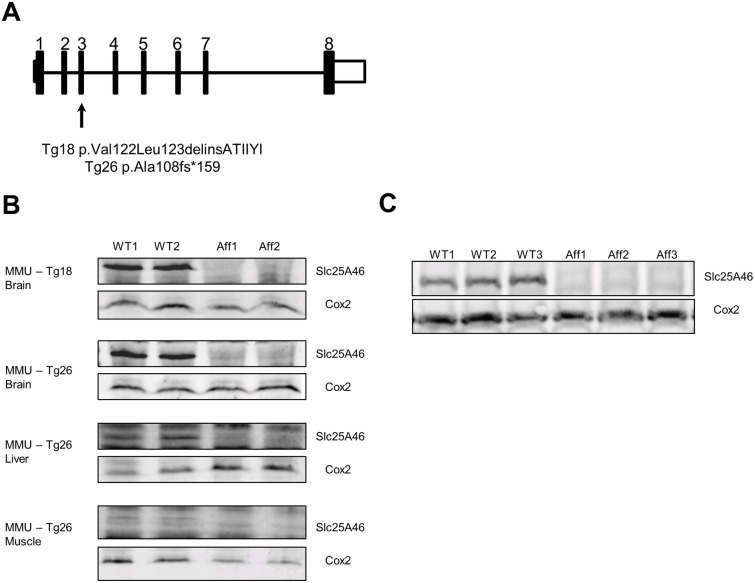
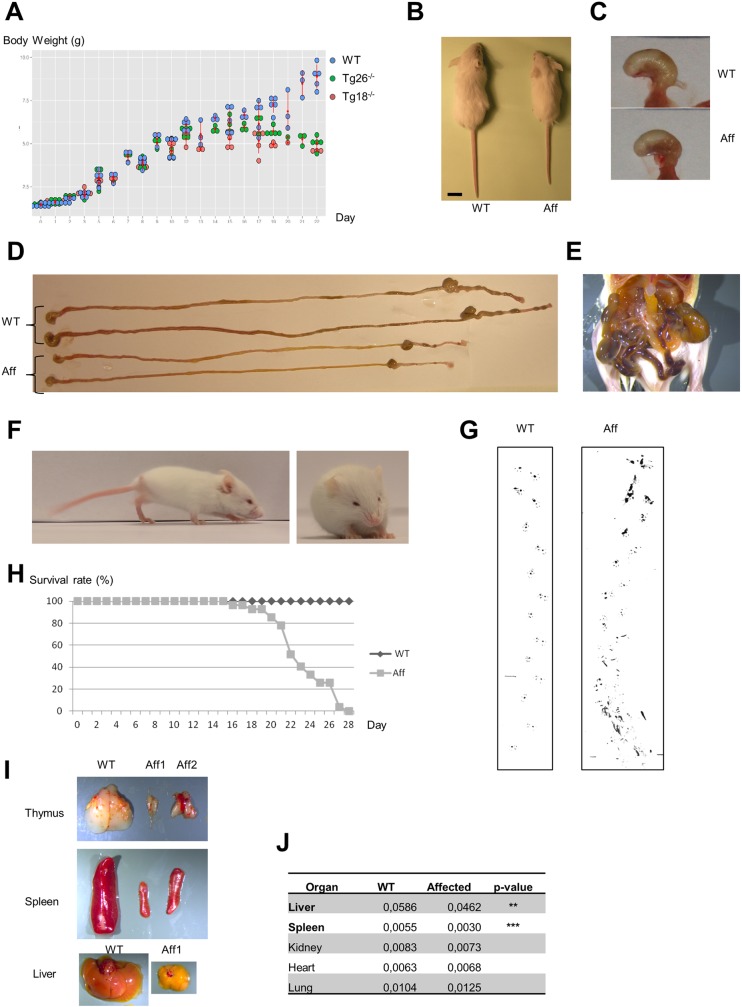
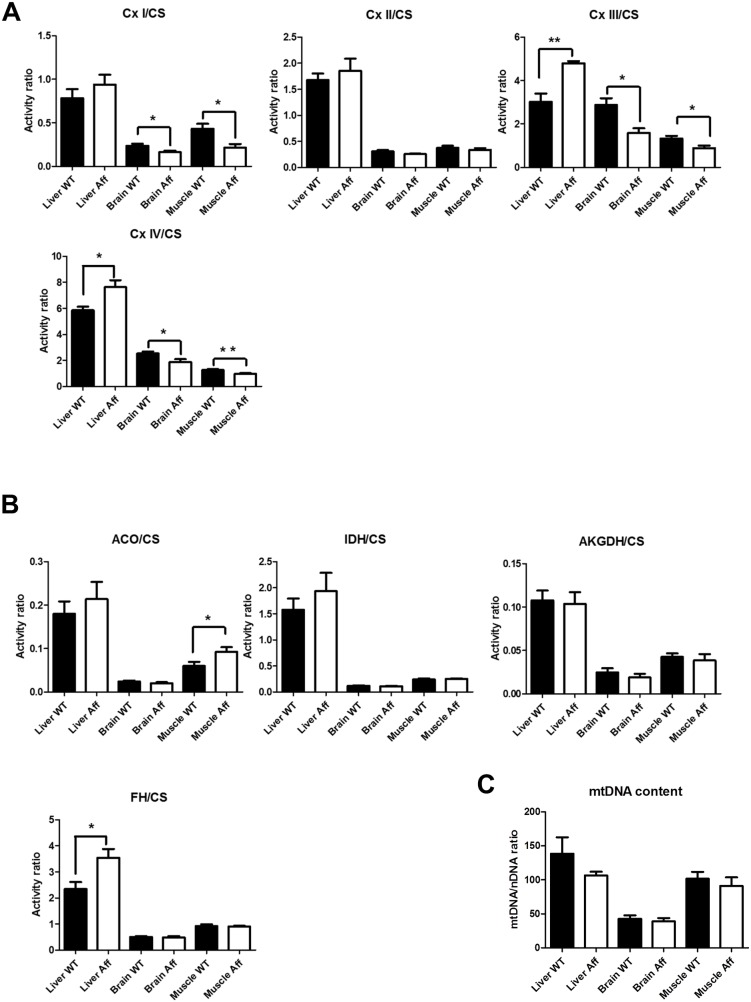
Neuropathies are neurodegenerative diseases affecting humans and other mammals. Many genetic causes have been identified so far, including mutations of genes encoding proteins involved in mitochondrial dynamics. Recently, the "Turning calves syndrome", a novel sensorimotor polyneuropathy was described in the French Rouge-des-Prés cattle breed. In the present study, we determined that this hereditary disease resulted from a single nucleotide substitution in SLC25A46, a gene encoding a protein of the mitochondrial carrier family. This mutation caused an apparent damaging amino-acid substitution. To better understand the function of this protein, we knocked out the Slc25a46 gene in a mouse model. This alteration affected not only the nervous system but also altered general metabolism, resulting in premature mortality. Based on optic microscopy examination, electron microscopy and on biochemical, metabolic and proteomic analyses, we showed that the Slc25a46 disruption caused a fusion/fission imbalance and an abnormal mitochondrial architecture that disturbed mitochondrial metabolism. These data extended the range of phenotypes associated with Slc25a46 dysfunction. Moreover, this Slc25a46 knock-out mouse model should be useful to further elucidate the role of SLC25A46 in mitochondrial dynamics.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
-
- Zick M, Rabl R, Reichert AS. Cristae formation-linking ultrastructure and function of mitochondria. Biochim Biophys Acta—Mol Cell Res [Internet]. Elsevier B.V.; 2009;1793(1):5–19. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
