

Bactericidal Permeability-Increasing Proteins Shape Host-Microbe Interactions
- PMID: 28377525
- PMCID: PMC5380838
- DOI: 10.1128/mBio.00040-17
Bactericidal Permeability-Increasing Proteins Shape Host-Microbe Interactions
Abstract
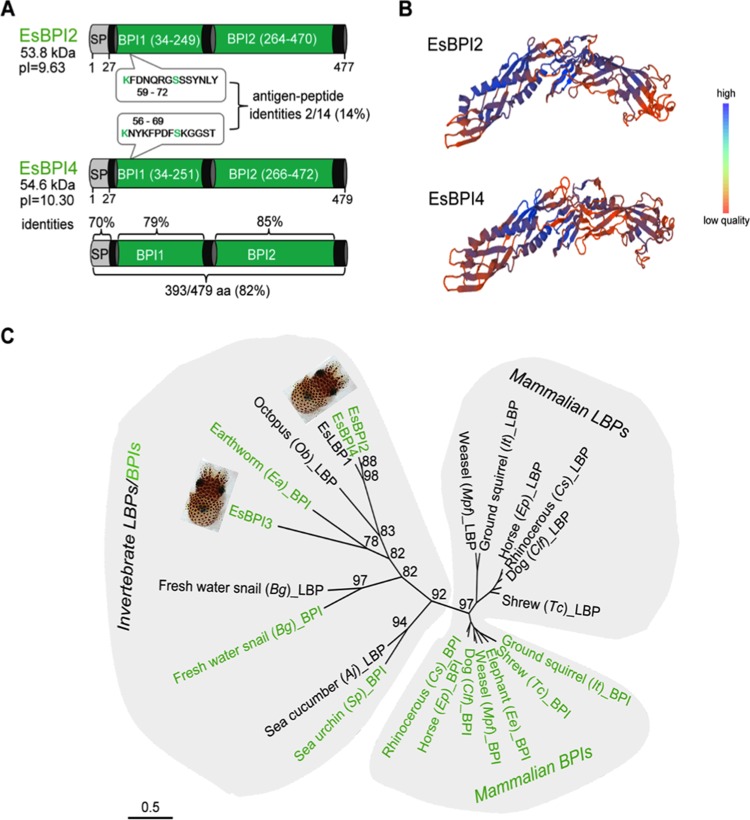
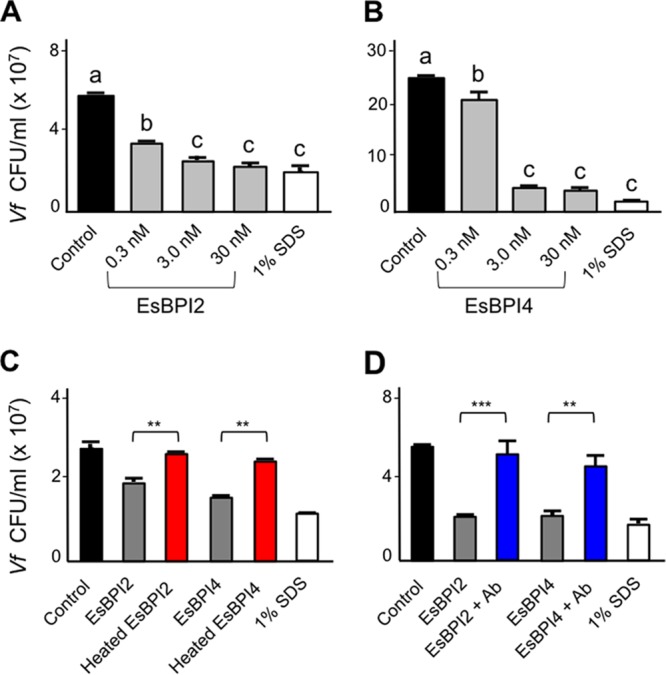
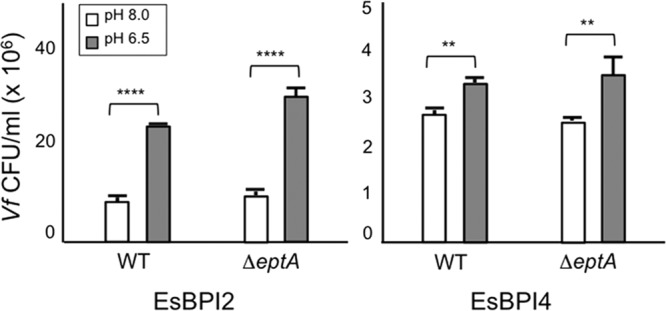
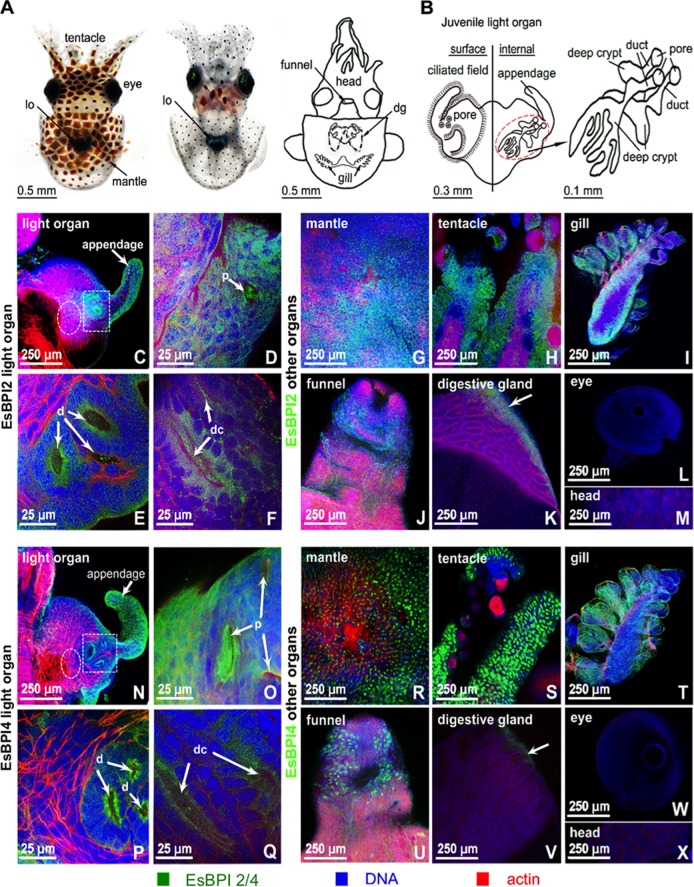
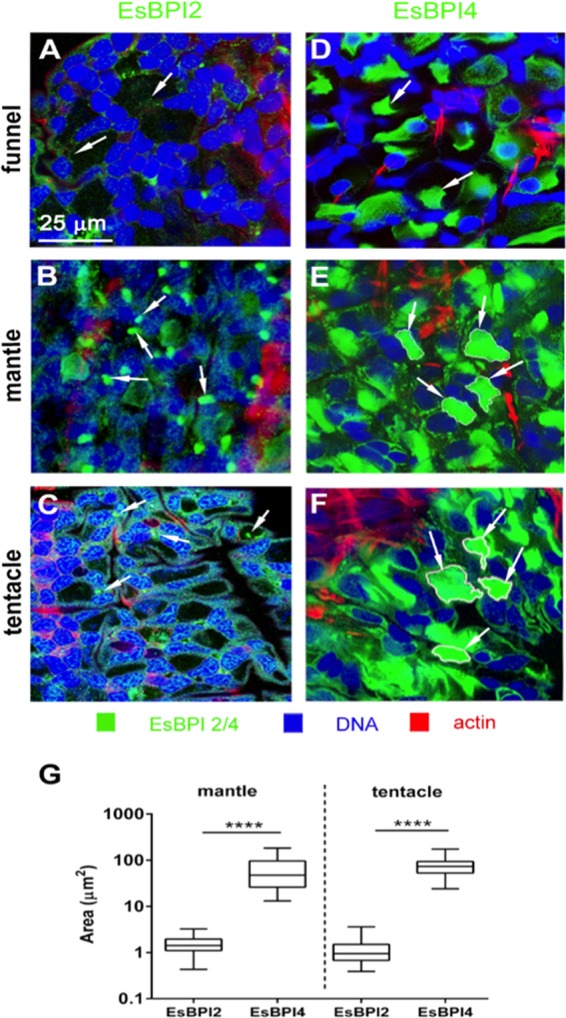
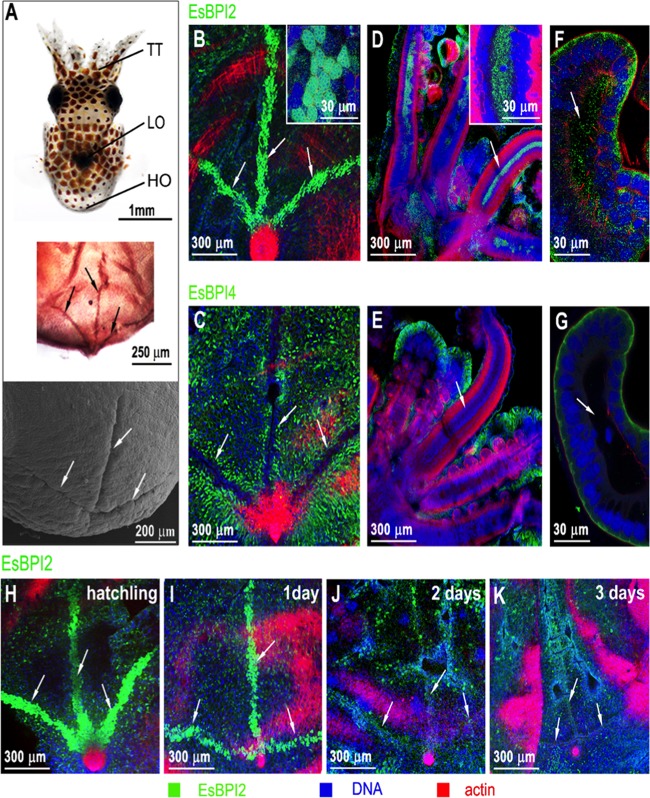
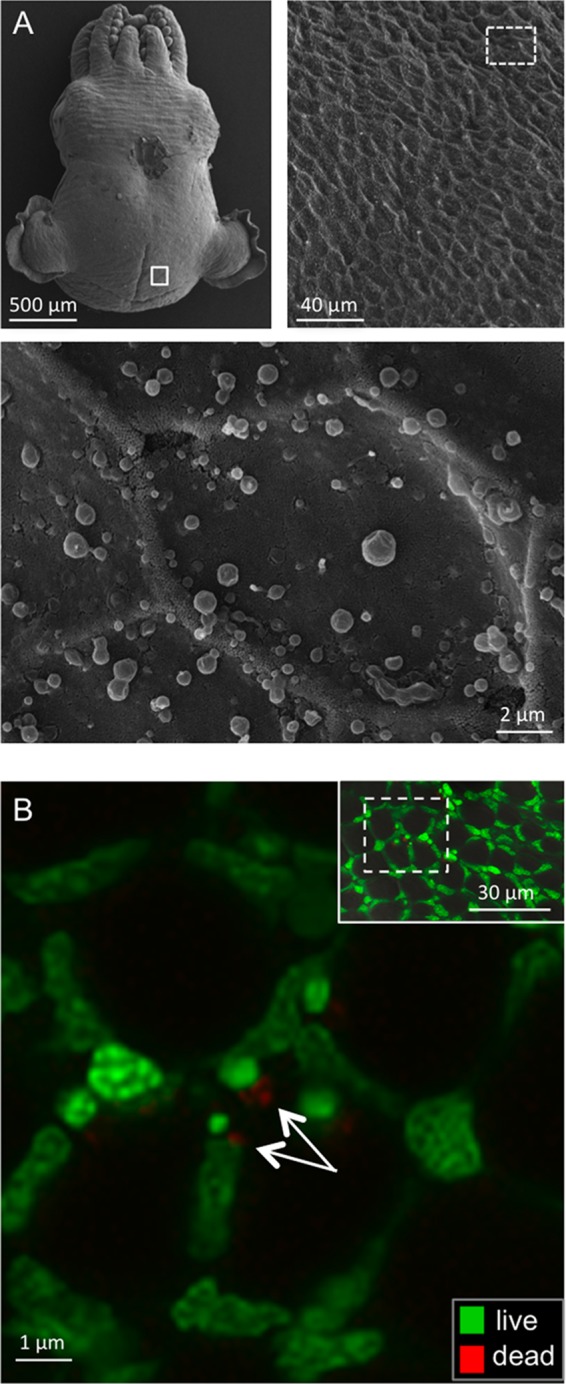
We characterized bactericidal permeability-increasing proteins (BPIs) of the squid Euprymna scolopes, EsBPI2 and EsBPI4. They have molecular characteristics typical of other animal BPIs, are closely related to one another, and nest phylogenetically among invertebrate BPIs. Purified EsBPIs had antimicrobial activity against the squid's symbiont, Vibrio fischeri, which colonizes light organ crypt epithelia. Activity of both proteins was abrogated by heat treatment and coincubation with specific antibodies. Pretreatment under acidic conditions similar to those during symbiosis initiation rendered V. fischeri more resistant to the antimicrobial activity of the proteins. Immunocytochemistry localized EsBPIs to the symbiotic organ and other epithelial surfaces interacting with ambient seawater. The proteins differed in intracellular distribution. Further, whereas EsBPI4 was restricted to epithelia, EsBPI2 also occurred in blood and in a transient juvenile organ that mediates hatching. The data provide evidence that these BPIs play different defensive roles early in the life of E. scolopes, modulating interactions with the symbiont.IMPORTANCE This study describes new functions for bactericidal permeability-increasing proteins (BPIs), members of the lipopolysaccharide-binding protein (LBP)/BPI protein family. The data provide evidence that these proteins play a dual role in the modulation of symbiotic bacteria. In the squid-vibrio model, these proteins both control the symbiont populations in the light organ tissues where symbiont cells occur in dense monoculture and, concomitantly, inhibit the symbiont from colonizing other epithelial surfaces of the animal.
Keywords: Vibrio fischeri; antimicrobial peptides; bioinformatics; confocal microscopy; symbiosis.
Copyright © 2017 Chen et al.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
