

The 5'UTR in human adenoviruses: leader diversity in late gene expression
- PMID: 28377580
- PMCID: PMC5429599
- DOI: 10.1038/s41598-017-00747-y
The 5'UTR in human adenoviruses: leader diversity in late gene expression
Abstract
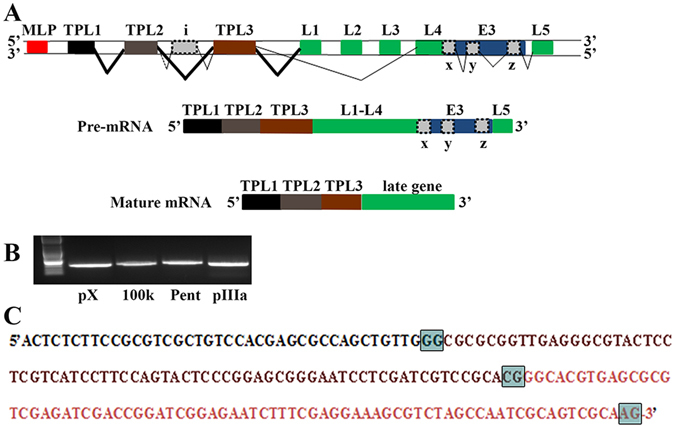
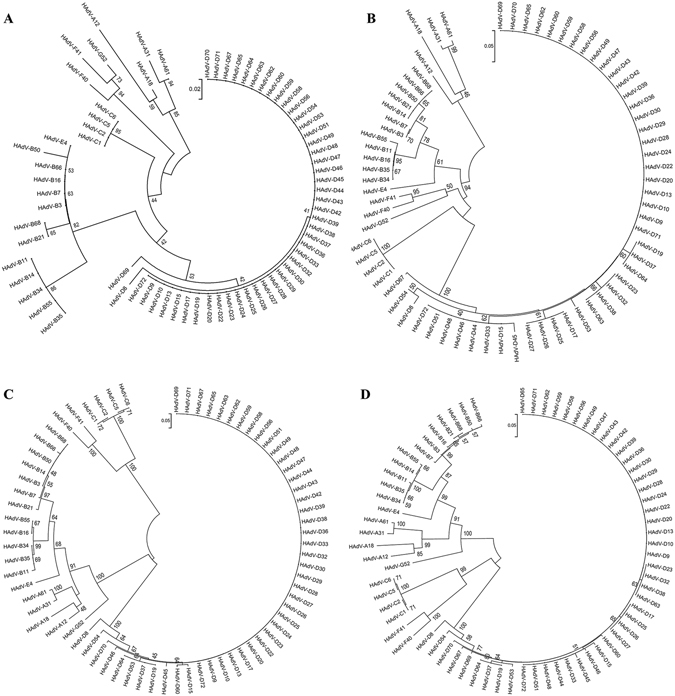
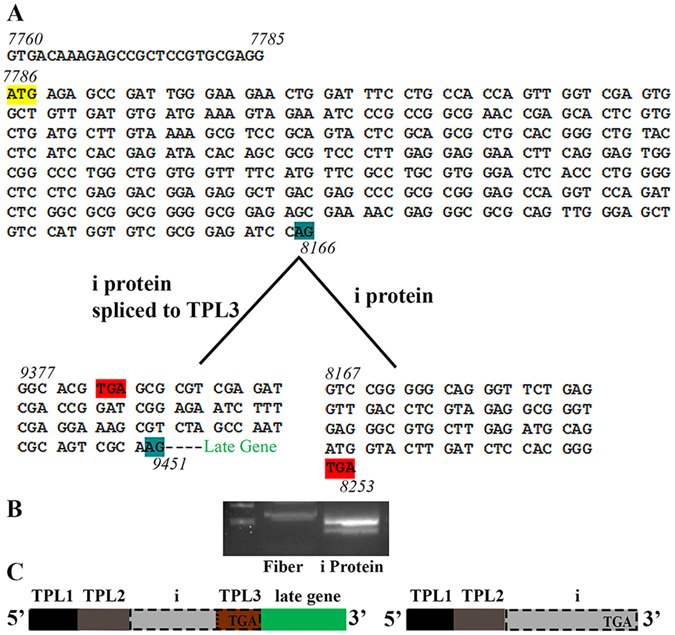
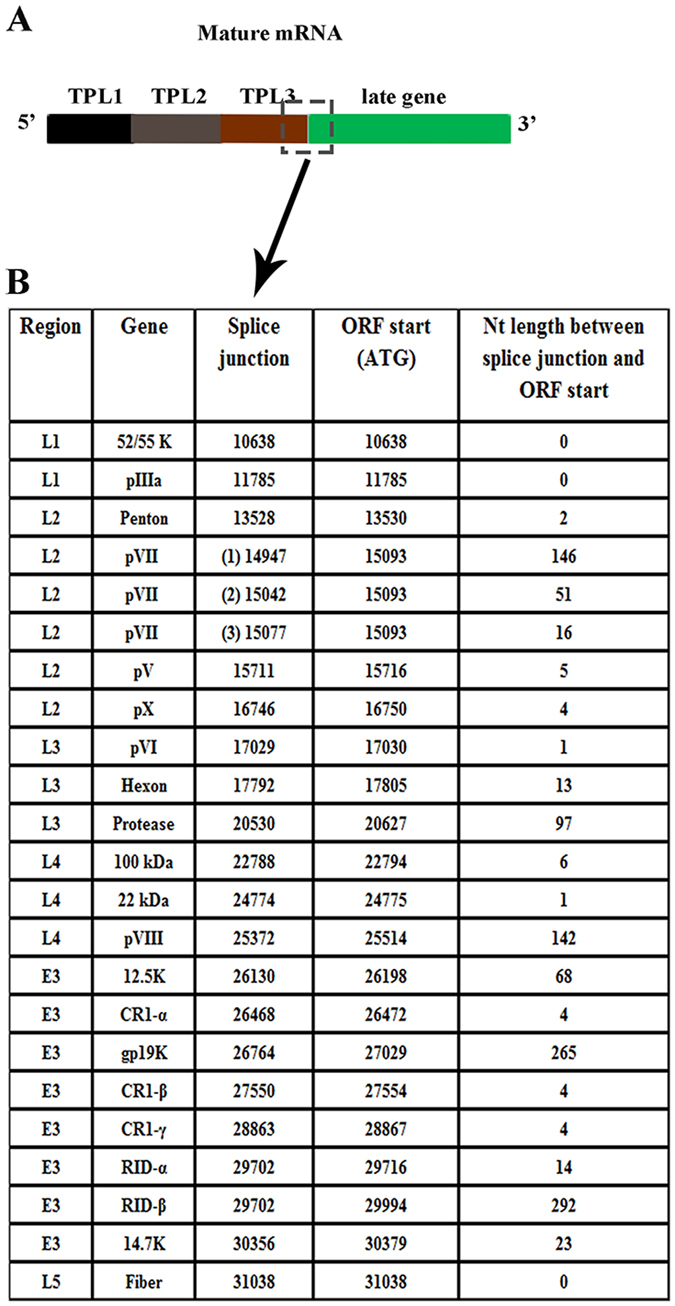
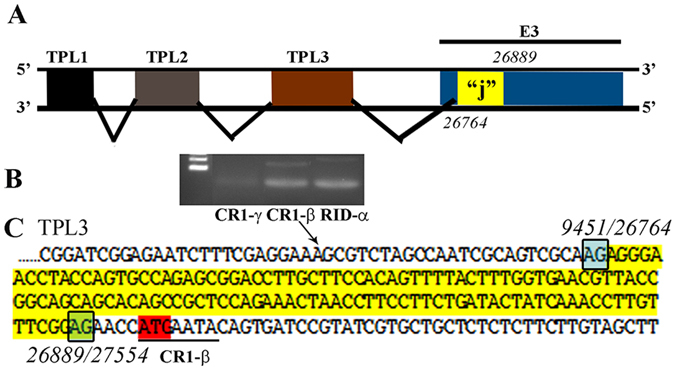
Human adenoviruses (HAdVs) shut down host cellular cap-dependent mRNA translation while initiating the translation of viral late mRNAs in a cap-independent manner. HAdV 5' untranslated regions (5'UTRs) are crucial for cap-independent initiation, and influence mRNA localization and stability. However, HAdV translational regulation remains relatively uncharacterized. The HAdV tripartite leader (TPL), composed of three introns (TPL 1-3), is critical to the translation of HAdV late mRNA. Herein, we annotated and analyzed 72 HAdV genotypes for the HAdV TPL and another previously described leader, the i-leader. Using HAdV species D, type 37 (HAdV-D37), we show by reverse transcription PCR and Sanger sequencing that mRNAs of the HAdV-D37 E3 transcription unit are spliced to the TPL. We also identified a polycistronic mRNA for RID-α and RID-β. Analysis of the i-leader revealed a potential open reading frame within the leader sequence and the termination of this potential protein in TPL3. A potential new leader embedded within the E3 region was also detected and tentatively named the j-leader. These results suggest an underappreciated complexity of post-transcriptional regulation, and the importance of HAdV 5'UTRs for precisely coordinated viral protein expression along the path from genotype to phenotype.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
