

Translation in astrocyte distal processes sets molecular heterogeneity at the gliovascular interface
- PMID: 28377822
- PMCID: PMC5368712
- DOI: 10.1038/celldisc.2017.5
Translation in astrocyte distal processes sets molecular heterogeneity at the gliovascular interface
Abstract
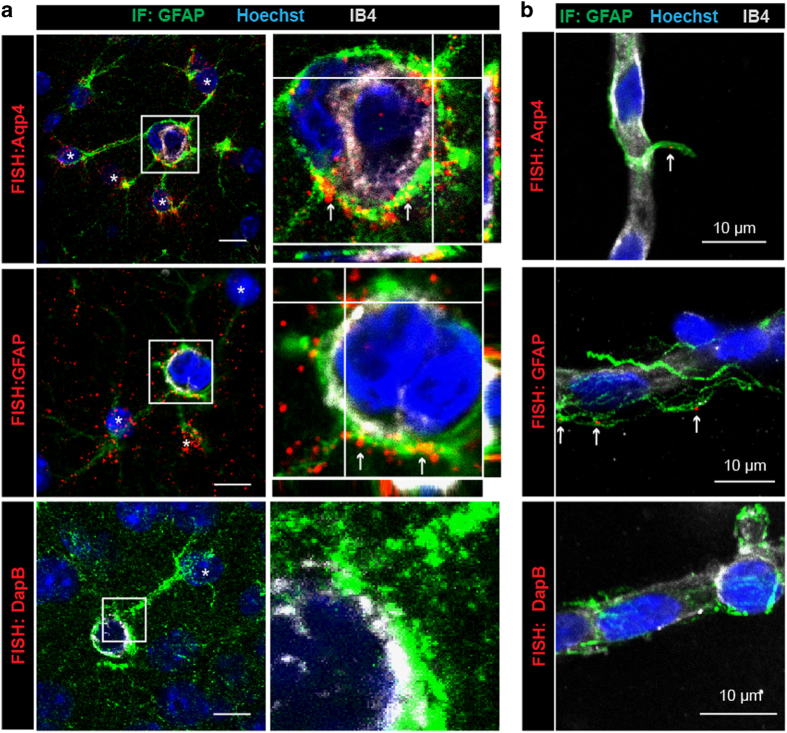
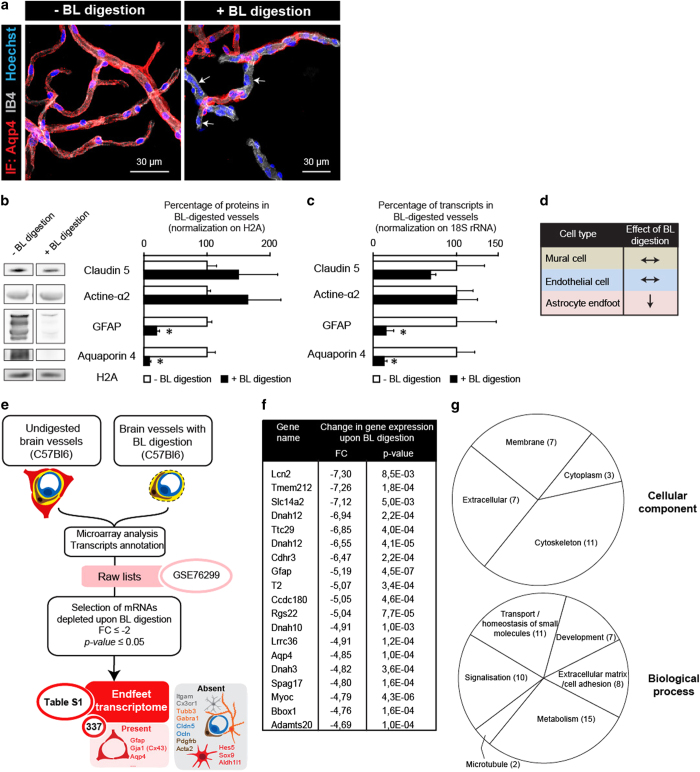
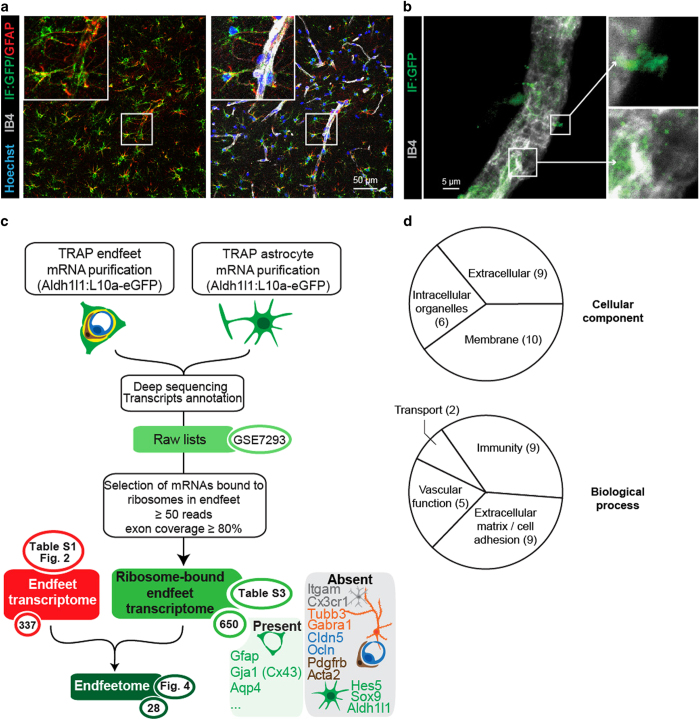
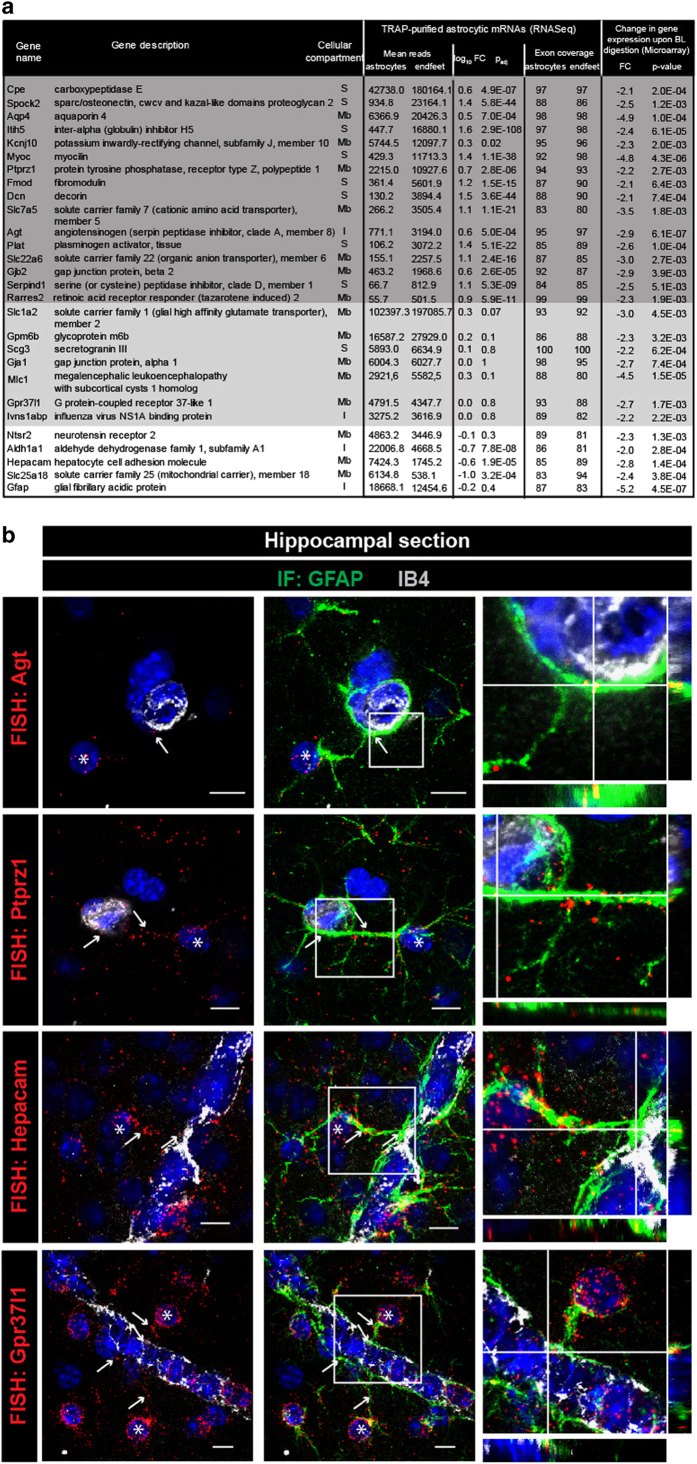
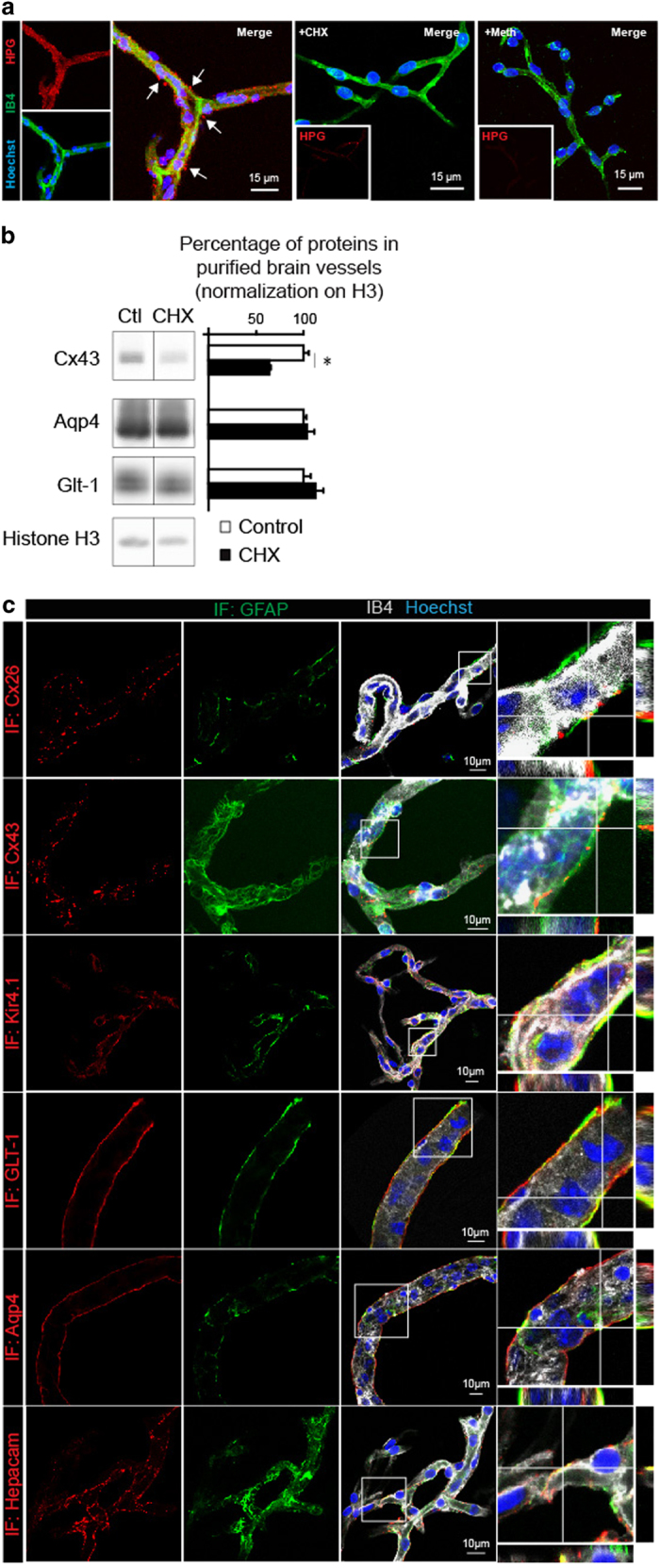
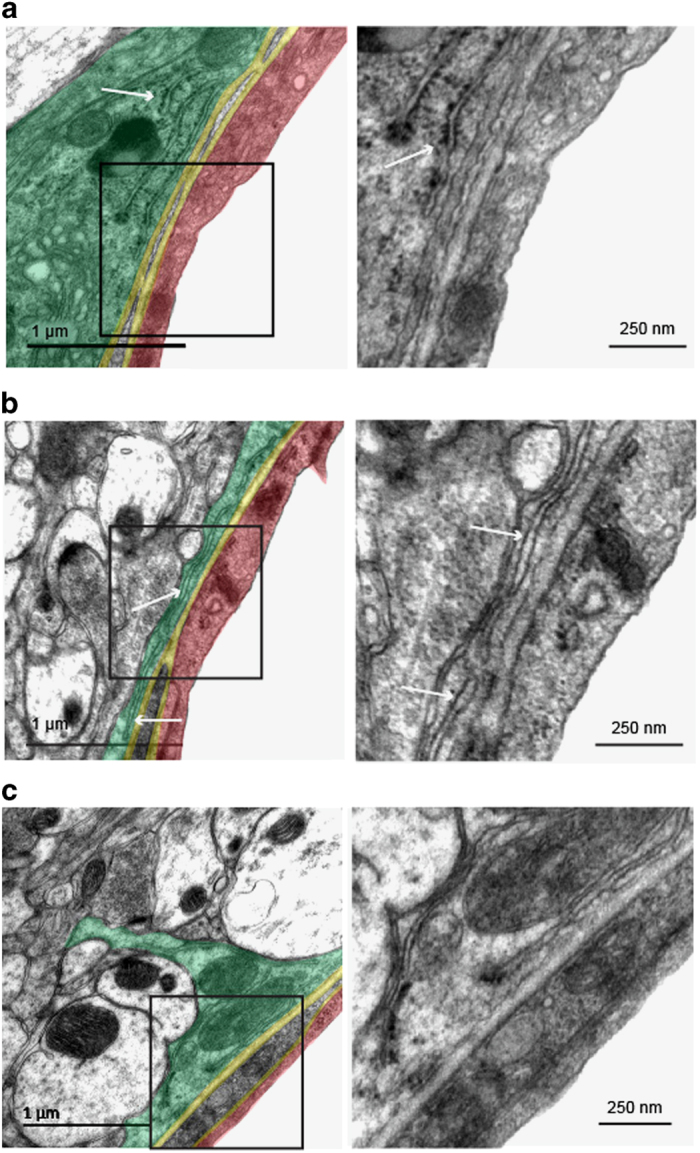
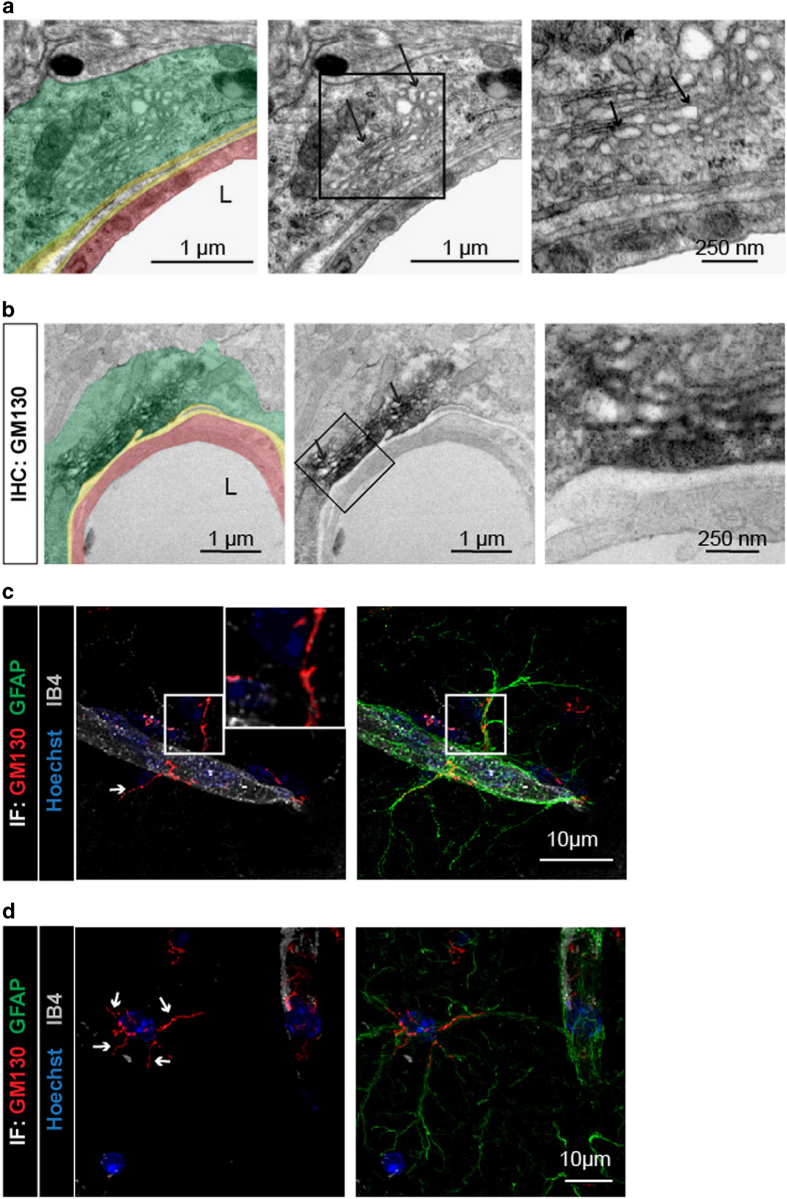
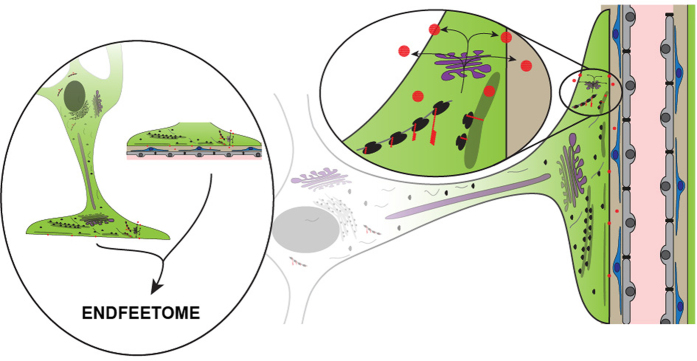
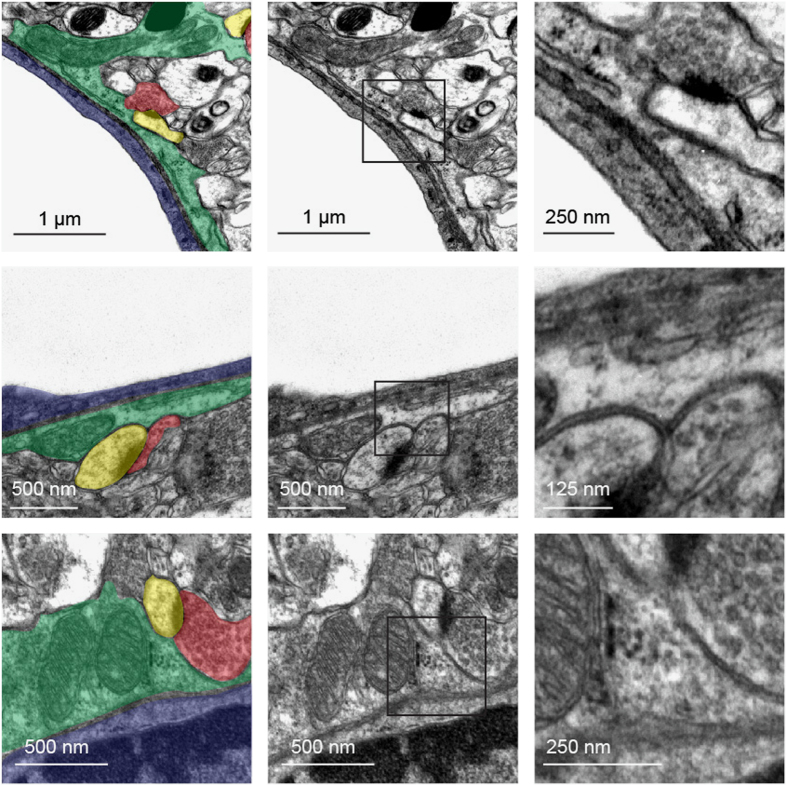
Astrocytes send out long processes that are terminated by endfeet at the vascular surface and regulate vascular functions as well as homeostasis at the vascular interface. To date, the astroglial mechanisms underlying these functions have been poorly addressed. Here we demonstrate that a subset of messenger RNAs is distributed in astrocyte endfeet. We identified, among this transcriptome, a pool of messenger RNAs bound to ribosomes, the endfeetome, that primarily encodes for secreted and membrane proteins. We detected nascent protein synthesis in astrocyte endfeet. Finally, we determined the presence of smooth and rough endoplasmic reticulum and the Golgi apparatus in astrocyte perivascular processes and endfeet, suggesting for local maturation of membrane and secreted proteins. These results demonstrate for the first time that protein synthesis occurs in astrocyte perivascular distal processes that may sustain their structural and functional polarization at the vascular interface.
Keywords: astrocyte; endfeet; gliovascular unit; local translation; mRNAs localization; translating ribosome immunoprecipitation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ghezali G, Dallerac G, Rouach N. Perisynaptic astroglial processes: dynamic processors of neuronal information. Brain Struct Funct 2015; 221: 2427–2442. - PubMed
-
- Mathiisen TM, Lehre KP, Danbolt NC, Ottersen OP. The perivascular astroglial sheath provides a complete covering of the brain microvessels: an electron microscopic 3D reconstruction. Glia 2010; 58: 1094–1103. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
