

Hepatic FXR/SHP axis modulates systemic glucose and fatty acid homeostasis in aged mice
- PMID: 28378930
- PMCID: PMC8156739
- DOI: 10.1002/hep.29199
Hepatic FXR/SHP axis modulates systemic glucose and fatty acid homeostasis in aged mice
Abstract
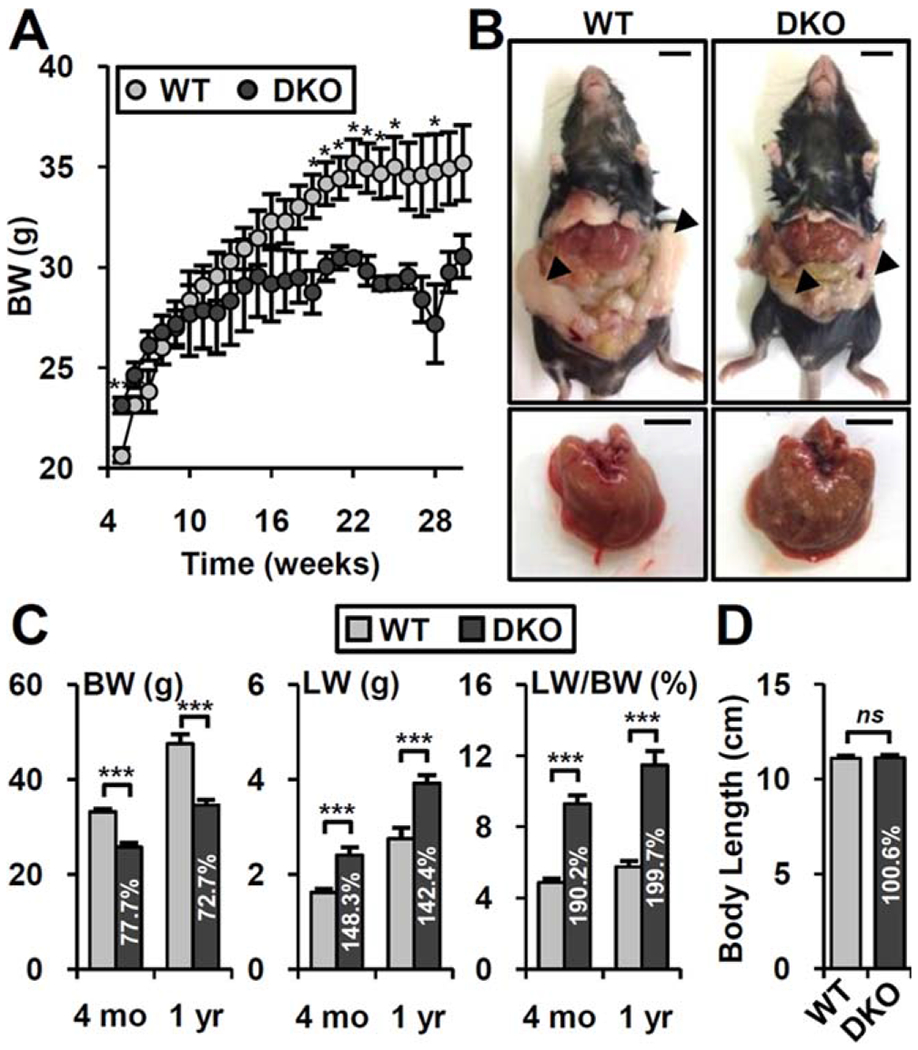
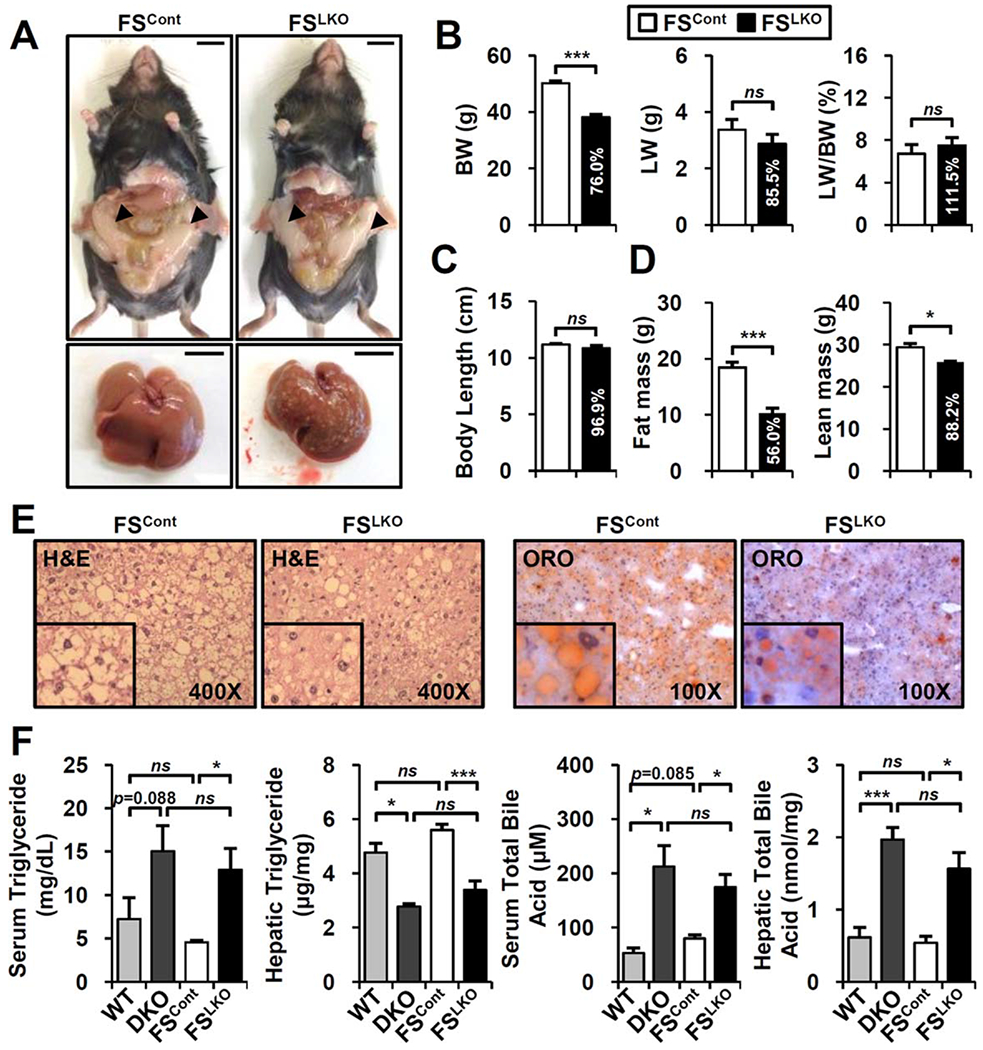
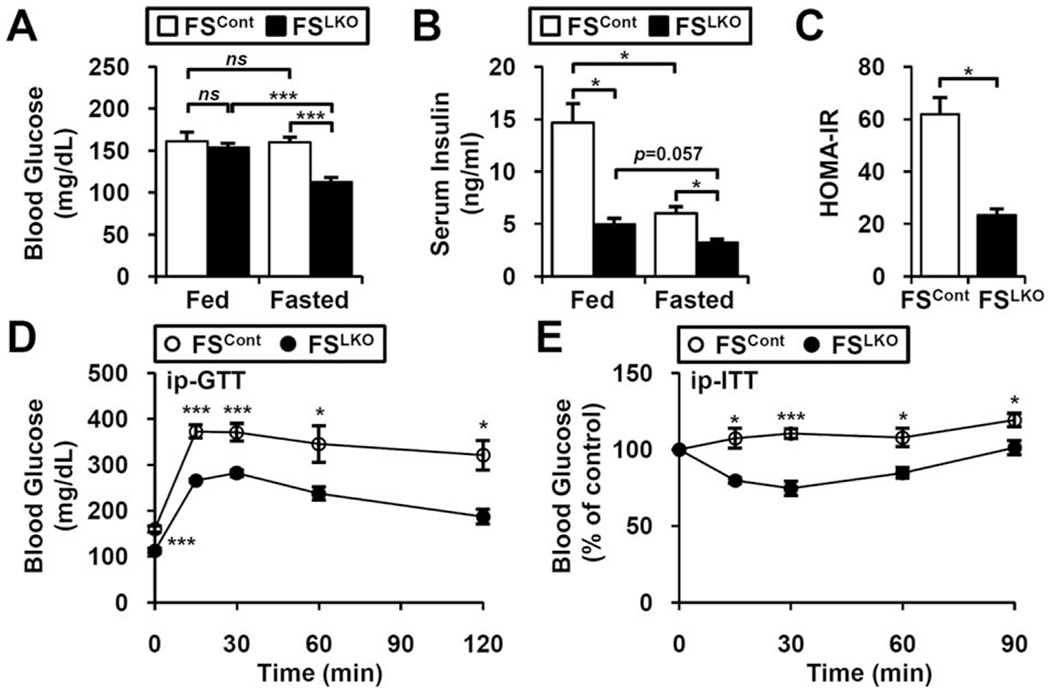
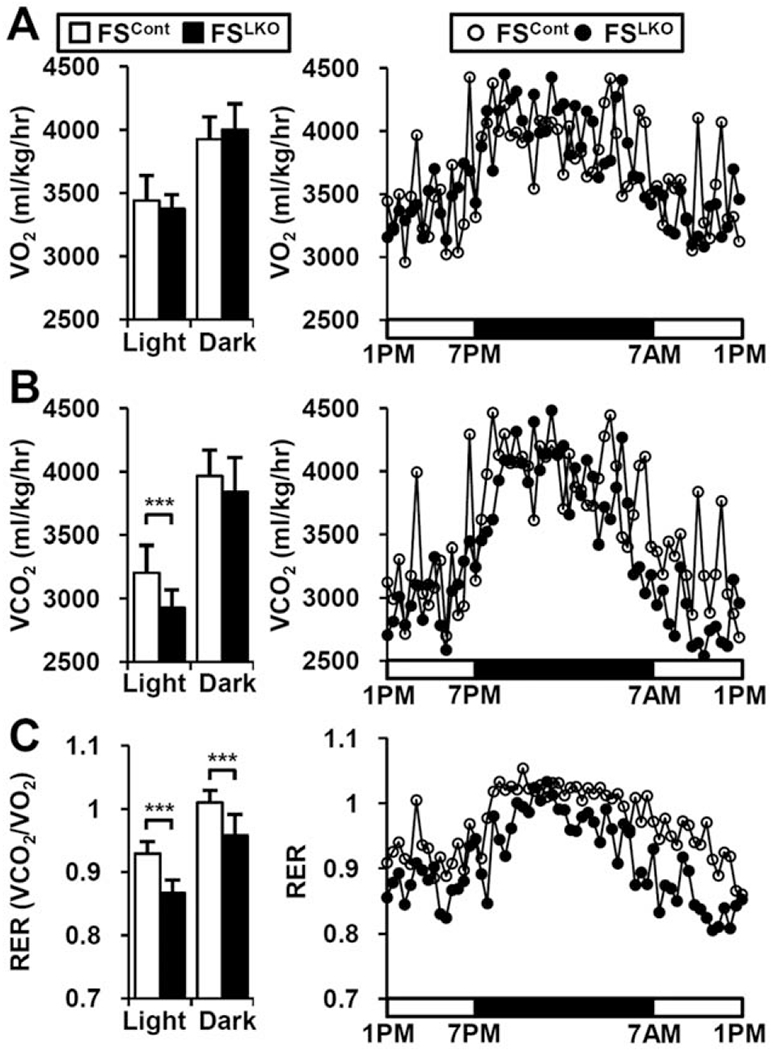
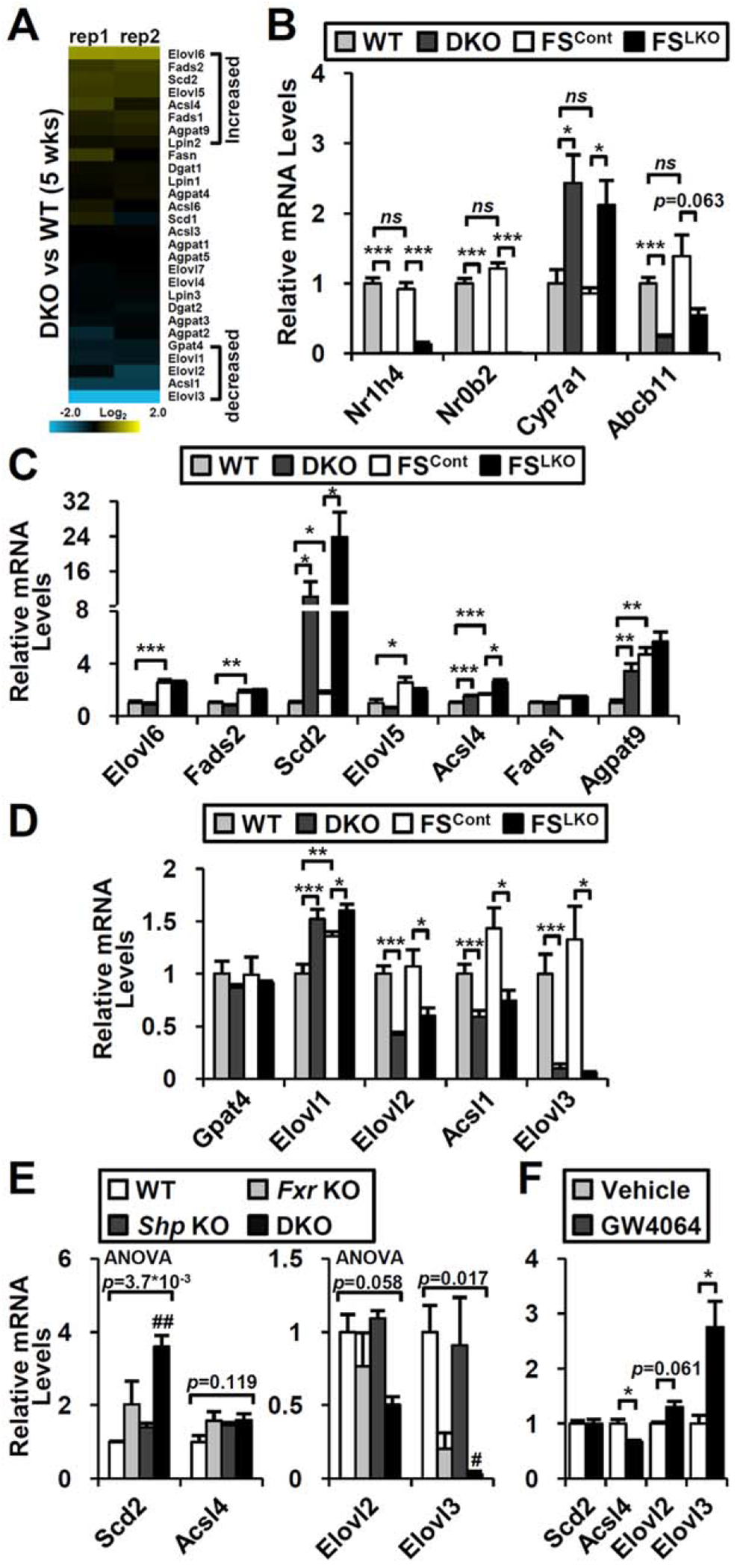
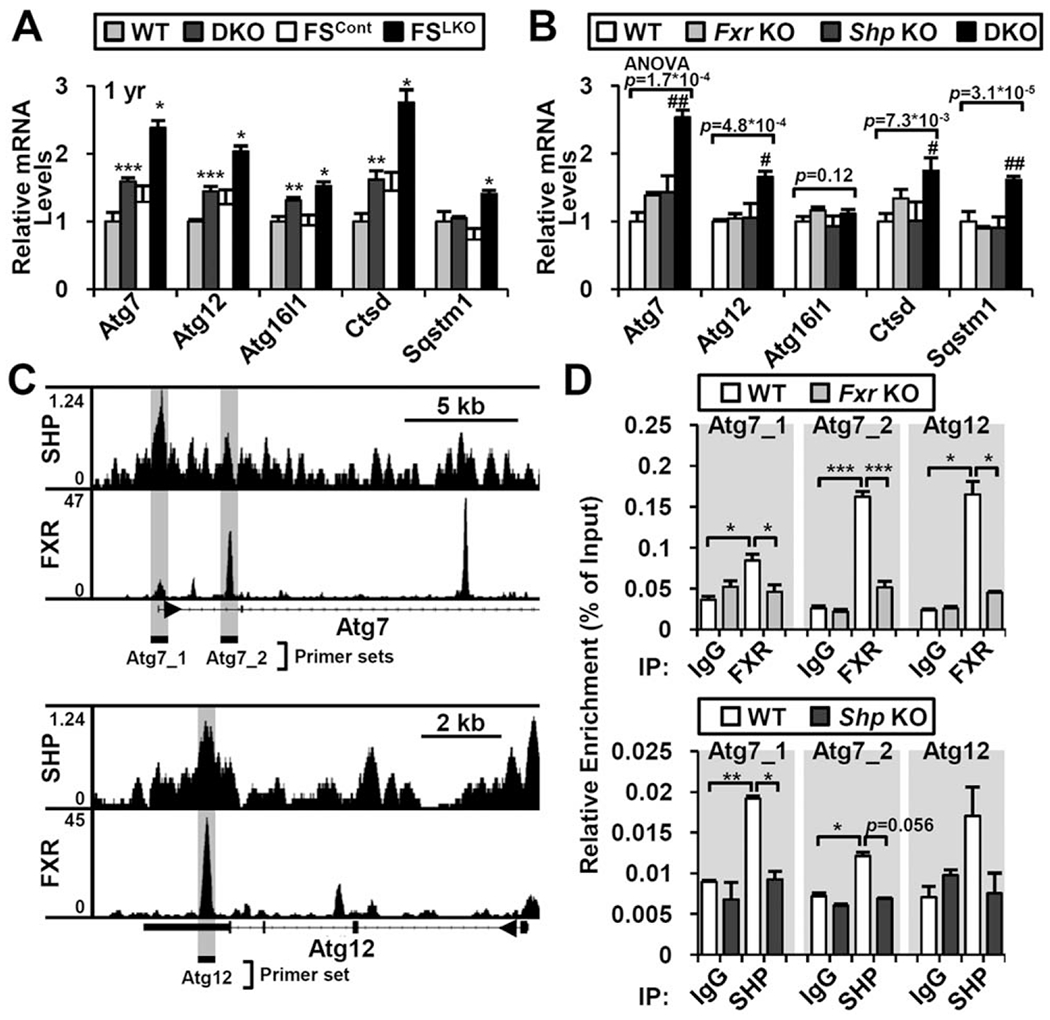
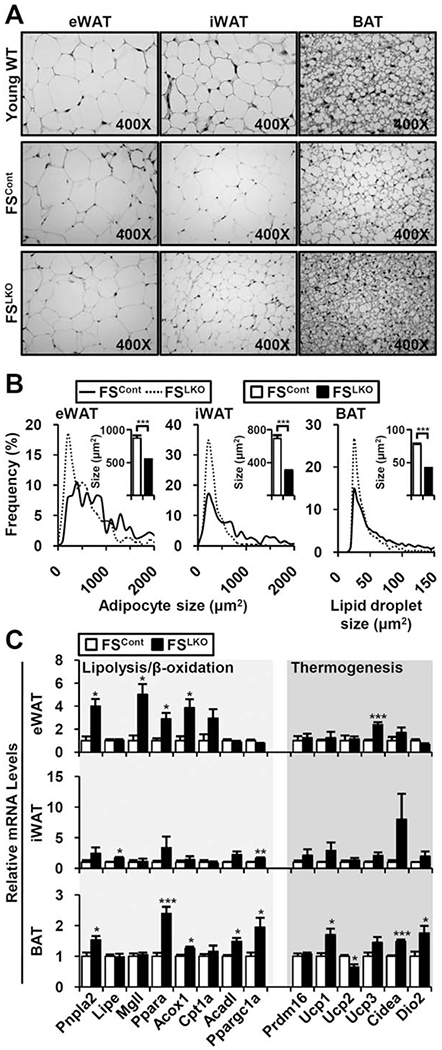
The nuclear receptors farnesoid X receptor (FXR; NR1H4) and small heterodimer partner (SHP; NR0B2) play crucial roles in bile acid homeostasis. Global double knockout of FXR and SHP signaling (DKO) causes severe cholestasis and liver injury at early ages. Here, we report an unexpected beneficial impact on glucose and fatty acid metabolism in aged DKO mice, which show suppressed body weight gain and adiposity when maintained on normal chow. This phenotype was not observed in single Fxr or Shp knockouts. Liver-specific Fxr/Shp double knockout mice fully phenocopied the DKO mice, with lower hepatic triglyceride accumulation, improved glucose/insulin tolerance, and accelerated fatty acid use. In both DKO and liver-specific Fxr/Shp double knockout livers, these metabolic phenotypes were associated with altered expression of fatty acid metabolism and autophagy-machinery genes. Loss of the hepatic FXR/SHP axis reprogrammed white and brown adipose tissue gene expression to boost fatty acid usage.
Conclusion: Combined deletion of the hepatic FXR/SHP axis improves glucose/fatty acid homeostasis in aged mice, reversing the aging phenotype of body weight gain, increased adiposity, and glucose/insulin tolerance, suggesting a central role of this axis in whole-body energy homeostasis. (Hepatology 2017;66:498-509).
© 2017 by the American Association for the Study of Liver Diseases.
Conflict of interest statement
Potential conflict of interest: Nothing to report.
Figures
References
-
- Fuchs CD, Traussnigg SA, Trauner M. Nuclear receptor modulation for the treatment of nonalcoholic fatty liver disease. Semin Liver Dis 2016;36:69–86. - PubMed
-
- Cariou B, van Harmelen K, Duran-Sandoval D, van Dijk TH, Grefhorst A, Abdelkarim M, et al. The farnesoid X receptor modulates adiposity and peripheral insulin sensitivity in mice. J Biol Chem 2006;281:11039–11049. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
