

Recreational concentrations of alcohol enhance synaptic inhibition of cerebellar unipolar brush cells via pre- and postsynaptic mechanisms
- PMID: 28381493
- PMCID: PMC5498730
- DOI: 10.1152/jn.00963.2016
Recreational concentrations of alcohol enhance synaptic inhibition of cerebellar unipolar brush cells via pre- and postsynaptic mechanisms
Abstract
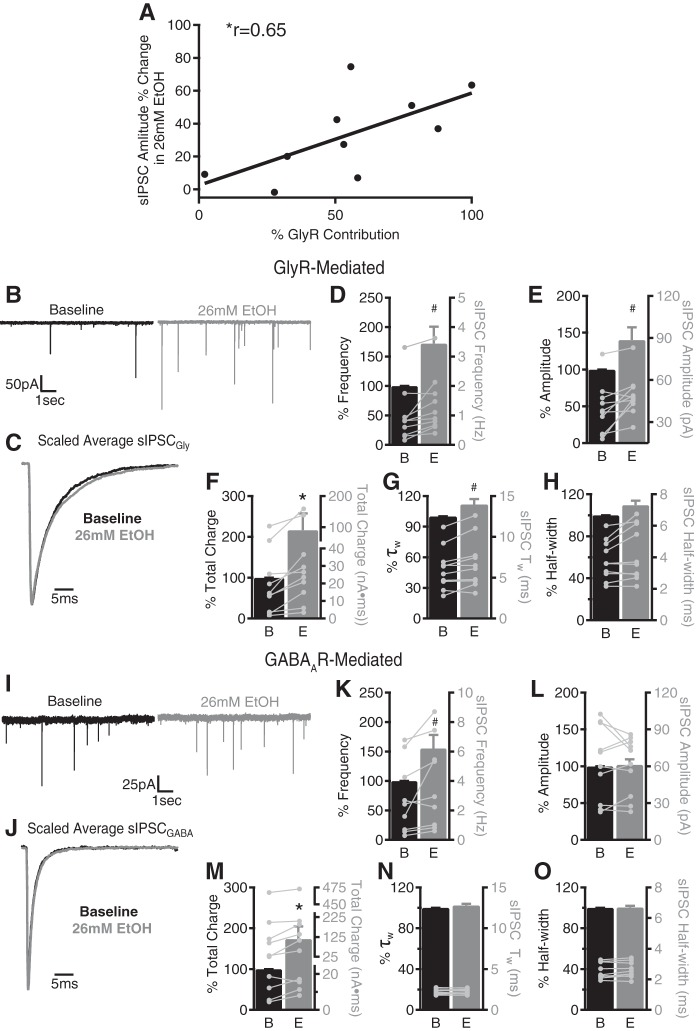
Variation in cerebellar sensitivity to alcohol/ethanol (EtOH) is a heritable trait associated with alcohol use disorder in humans and high EtOH consumption in rodents, but the underlying mechanisms are poorly understood. A recently identified cellular substrate of cerebellar sensitivity to EtOH, the GABAergic system of cerebellar granule cells (GCs), shows divergent responses to EtOH paralleling EtOH consumption and motor impairment phenotype. Although GCs are the dominant afferent integrator in the cerebellum, such integration is shared by unipolar brush cells (UBCs) in vestibulocerebellar lobes. UBCs receive both GABAergic and glycinergic inhibition, both of which may mediate diverse neurological effects of EtOH. Therefore, the impact of recreational concentrations of EtOH (~10-50 mM) on GABAA receptor (GABAAR)- and glycine receptor (GlyR)-mediated spontaneous inhibitory postsynaptic currents (sIPSCs) of UBCs in cerebellar slices was characterized. Sprague-Dawley rat (SDR) UBCs exhibited sIPSCs mediated by GABAARs, GlyRs, or both, and EtOH dose-dependently (10, 26, 52 mM) increased their frequency and amplitude. EtOH increased the frequency of glycinergic and GABAergic sIPSCs and selectively enhanced the amplitude of glycinergic sIPSCs. This GlyR-specific enhancement of sIPSC amplitude resulted from EtOH actions at presynaptic Golgi cells and via protein kinase C-dependent direct actions on postsynaptic GlyRs. The magnitude of EtOH-induced increases in UBC sIPSC activity varied across SDRs and two lines of mice, in parallel with their respective alcohol consumption/motor impairment phenotypes. These data indicate that Golgi cell-to-UBC inhibitory synapses are targets of EtOH, which acts at pre- and postsynaptic sites, via Golgi cell excitation and direct GlyR enhancement.NEW & NOTEWORTHY Genetic variability in cerebellar alcohol/ethanol sensitivity (ethanol-induced ataxia) predicts ethanol consumption phenotype in rodents and humans, but the cellular and molecular mechanisms underlying genetic differences are largely unknown. Here it is demonstrated that recreational concentrations of alcohol (10-30 mM) enhance glycinergic and GABAergic inhibition of unipolar brush cells through increases in glycine/GABA release and postsynaptic enhancement of glycine receptor-mediated responses. Ethanol effects varied across rodent genotypes parallel to ethanol consumption and motor sensitivity phenotype.
Keywords: alcohol; cerebellum; unipolar brush cell.
Copyright © 2017 the American Physiological Society.
Figures






References
-
- Aguayo LG, Pancetti FC. Ethanol modulation of the gamma-aminobutyric acidA- and glycine-activated Cl− current in cultured mouse neurons. J Pharmacol Exp Ther 270: 61–69, 1994. - PubMed
-
- Aguayo LG, Tapia JC, Pancetti FC. Potentiation of the glycine-activated Cl− current by ethanol in cultured mouse spinal neurons. J Pharmacol Exp Ther 279: 1116–1122, 1996. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous