

Essential role for non-canonical poly(A) polymerase GLD4 in cytoplasmic polyadenylation and carbohydrate metabolism
- PMID: 28383716
- PMCID: PMC5499868
- DOI: 10.1093/nar/gkx239
Essential role for non-canonical poly(A) polymerase GLD4 in cytoplasmic polyadenylation and carbohydrate metabolism
Abstract
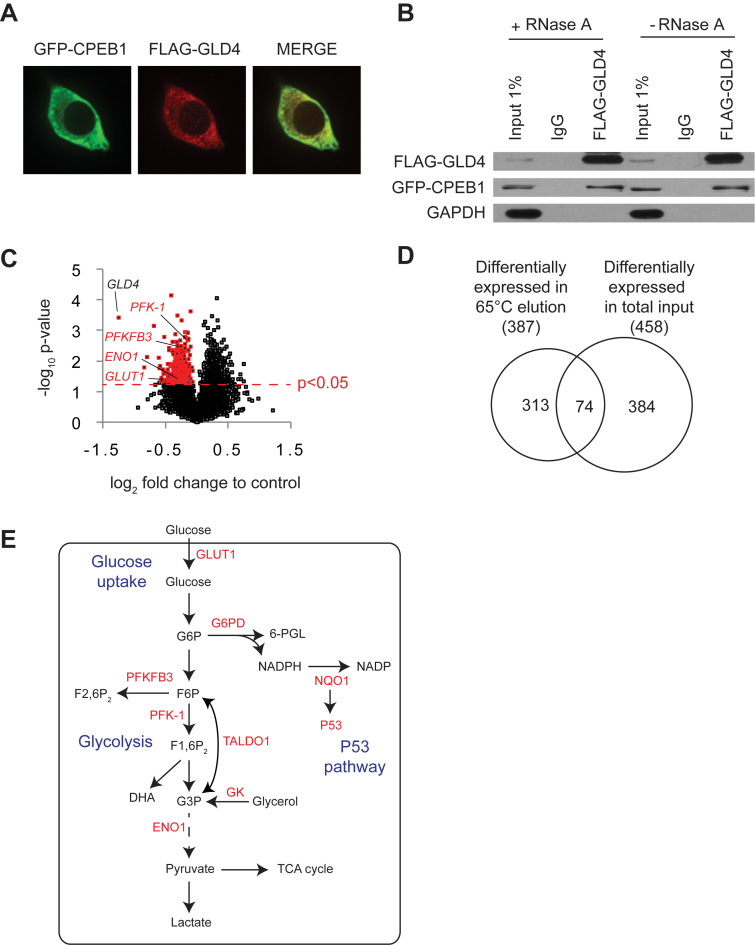
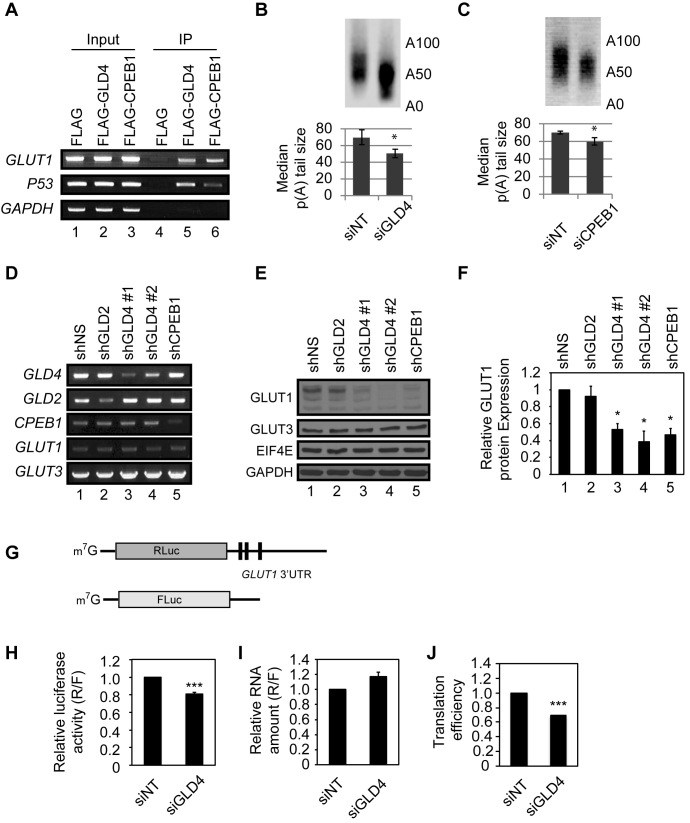
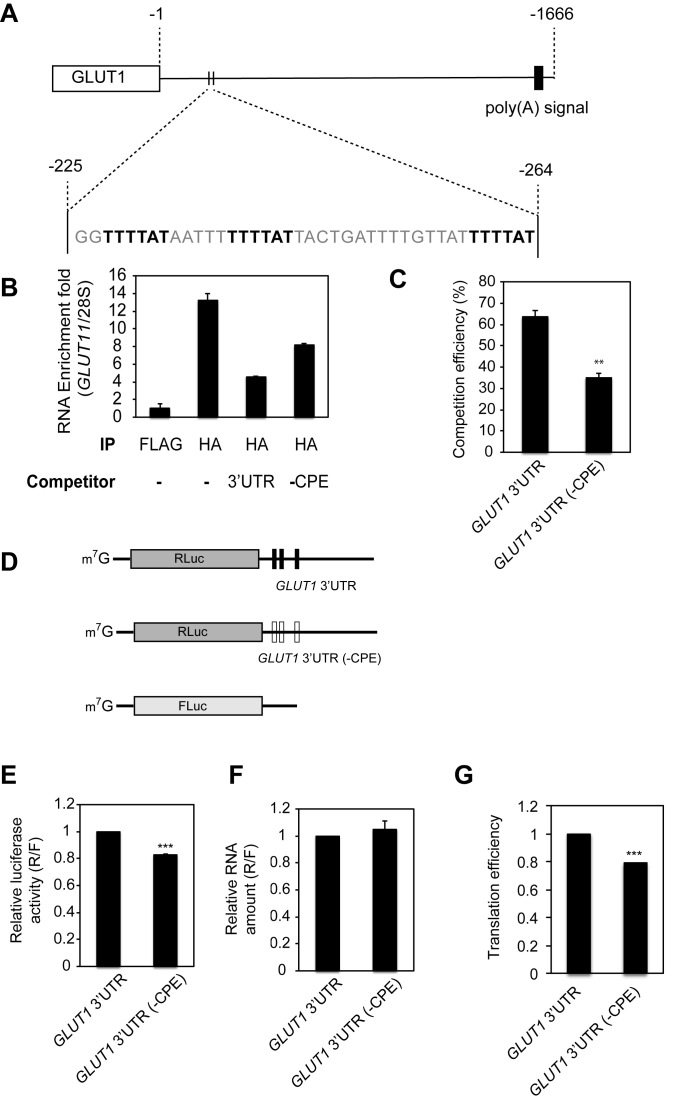
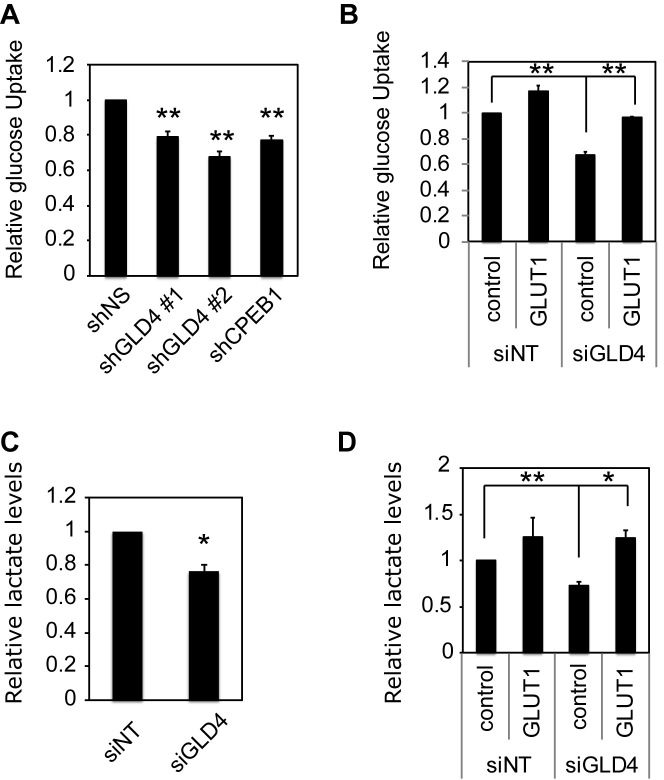
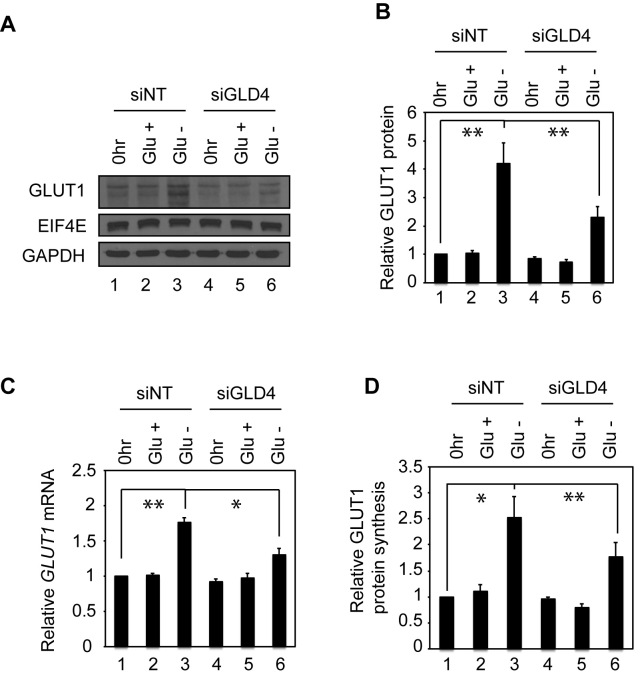
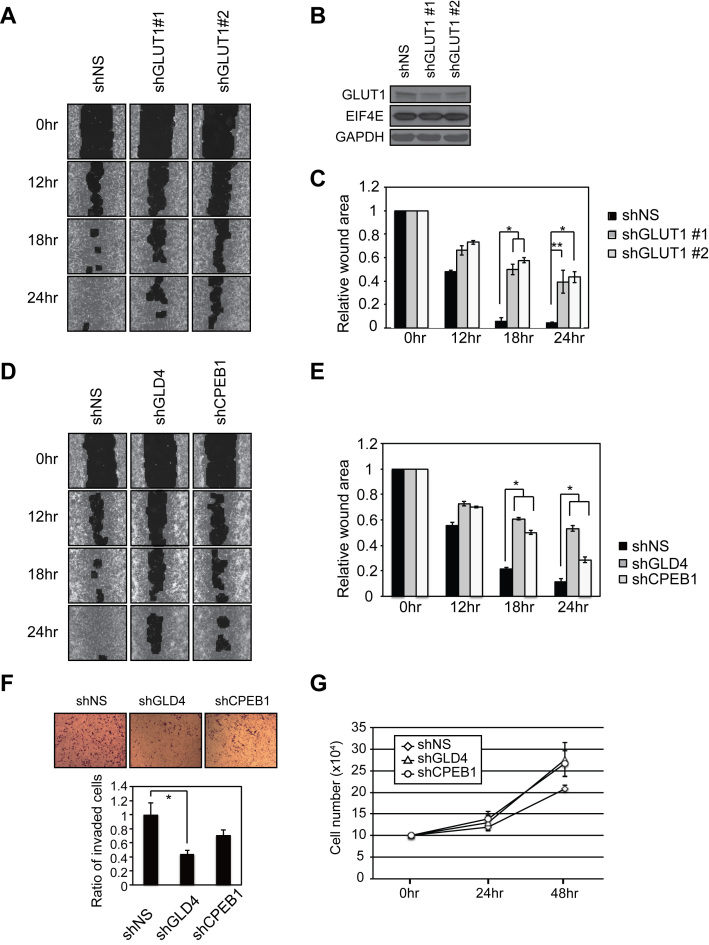
Regulation of gene expression at the level of cytoplasmic polyadenylation is important for many biological phenomena including cell cycle progression, mitochondrial respiration, and learning and memory. GLD4 is one of the non-canonical poly(A) polymerases that regulates cytoplasmic polyadenylation-induced translation, but its target mRNAs and role in cellular physiology is not well known. To assess the full panoply of mRNAs whose polyadenylation is controlled by GLD4, we performed an unbiased whole genome-wide screen using poy(U) chromatography and thermal elution. We identified hundreds of mRNAs regulated by GLD4, several of which are involved in carbohydrate metabolism including GLUT1, a major glucose transporter. Depletion of GLD4 not only reduced GLUT1 poly(A) tail length, but also GLUT1 protein. GLD4-mediated translational control of GLUT1 mRNA is dependent of an RNA binding protein, CPEB1, and its binding elements in the 3΄ UTR. Through regulating GLUT1 level, GLD4 affects glucose uptake into cells and lactate levels. Moreover, GLD4 depletion impairs glucose deprivation-induced GLUT1 up-regulation. In addition, we found that GLD4 affects glucose-dependent cellular phenotypes such as migration and invasion in glioblastoma cells. Our observations delineate a novel post-transcriptional regulatory network involving carbohydrate metabolism and glucose homeostasis mediated by GLD4.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Schmidt M.J., Norbury C.J.. Polyadenylation and beyond: emerging roles for noncanonical poly(A) polymerases. Wiley Interdiscipl. Rev. RNA. 2010; 1:142–151. - PubMed
-
- Mellman D.L., Gonzales M.L., Song C., Barlow C.A., Wang P., Kendziorski C., Anderson R.A.. A PtdIns4,5P2-regulated nuclear poly(A) polymerase controls expression of select mRNAs. Nature. 2008; 451:1013–1017. - PubMed
-
- Barnard D.C., Ryan K., Manley J.L., Richter J.D.. Symplekin and xGLD-2 are required for CPEB-mediated cytoplasmic polyadenylation. Cell. 2004; 119:641–651. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
