

Who lives in a fungus? The diversity, origins and functions of fungal endobacteria living in Mucoromycota
- PMID: 28387771
- PMCID: PMC5520026
- DOI: 10.1038/ismej.2017.21
Who lives in a fungus? The diversity, origins and functions of fungal endobacteria living in Mucoromycota
Abstract
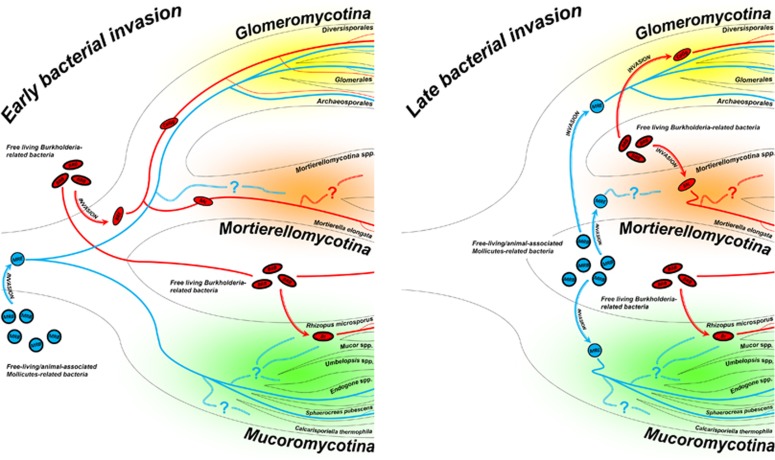
Bacterial interactions with plants and animals have been examined for many years; differently, only with the new millennium the study of bacterial-fungal interactions blossomed, becoming a new field of microbiology with relevance to microbial ecology, human health and biotechnology. Bacteria and fungi interact at different levels and bacterial endosymbionts, which dwell inside fungal cells, provide the most intimate example. Bacterial endosymbionts mostly occur in fungi of the phylum Mucoromycota and include Betaproteobacteria (Burkhoderia-related) and Mollicutes (Mycoplasma-related). Based on phylogenomics and estimations of divergence time, we hypothesized two different scenarios for the origin of these interactions (early vs late bacterial invasion). Sequencing of the genomes of fungal endobacteria revealed a significant reduction in genome size, particularly in endosymbionts of Glomeromycotina, as expected by their uncultivability and host dependency. Similar to endobacteria of insects, the endobacteria of fungi show a range of behaviours from mutualism to antagonism. Emerging results suggest that some benefits given by the endobacteria to their plant-associated fungal host may propagate to the interacting plant, giving rise to a three-level inter-domain interaction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ash C, Mueller K. (2016). Manipulating the microbiota. Science 352: 530–531. - PubMed
-
- Bianciotto V, Lumini E, Bonfante P, Vandamme P. (2003). 'Candidatus Glomeribacter gigasporarum' gen. nov., sp. nov., an endosymbiont of arbuscular mycorrhizal fungi. Int Syst Evol Microbiol 53: 121–124. - PubMed
-
- Bonfante P, Scannerini S. (1977). Cytological observations on the mycorrhiza Endogone flammicorona-Pinus strobus. Allionia 22: 23–34.
-
- Bonfante P, Scannerini S. (1976). The ultrastructure of the zygospore in Endogone flammicorona Trappe and Gerdemann. Mycopathologia 59: 117–123.
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
