

Development of a glycoconjugate vaccine to prevent invasive Salmonella Typhimurium infections in sub-Saharan Africa
- PMID: 28388624
- PMCID: PMC5397072
- DOI: 10.1371/journal.pntd.0005493
Development of a glycoconjugate vaccine to prevent invasive Salmonella Typhimurium infections in sub-Saharan Africa
Abstract
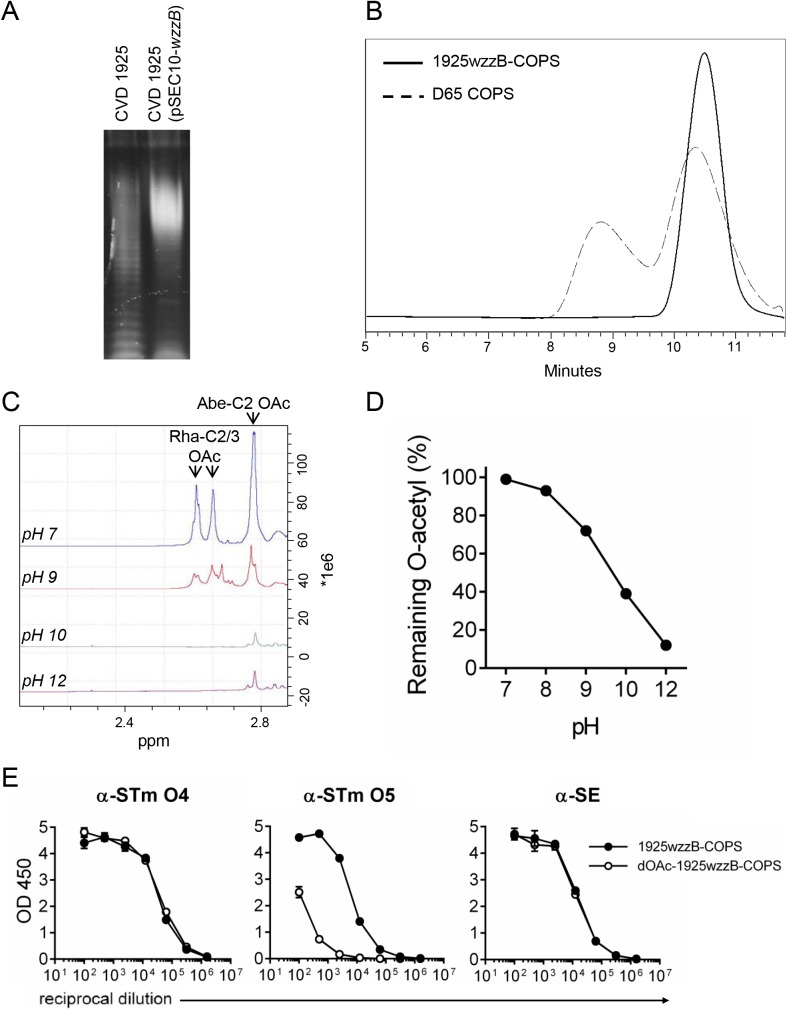
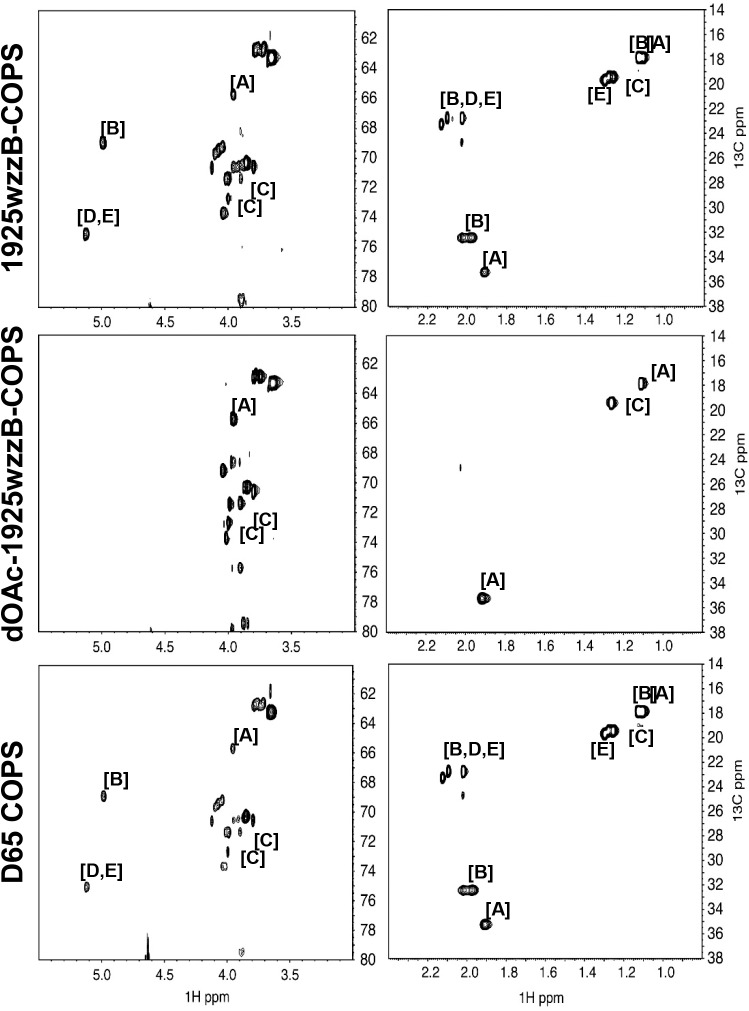
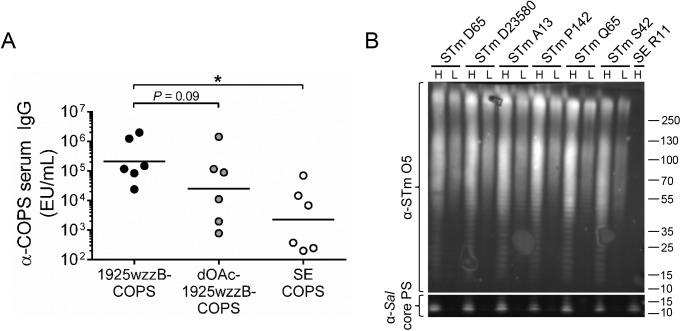
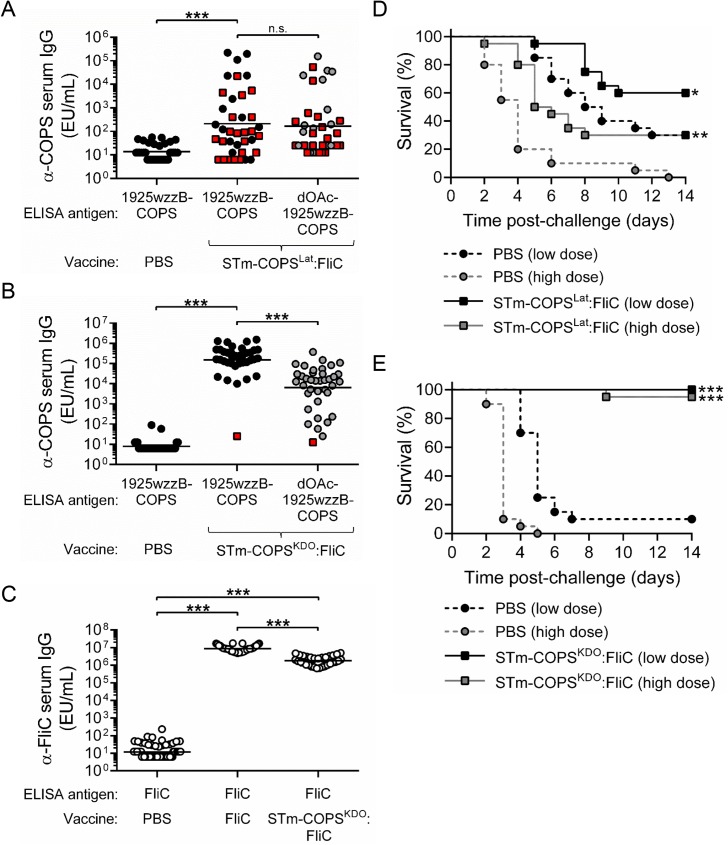
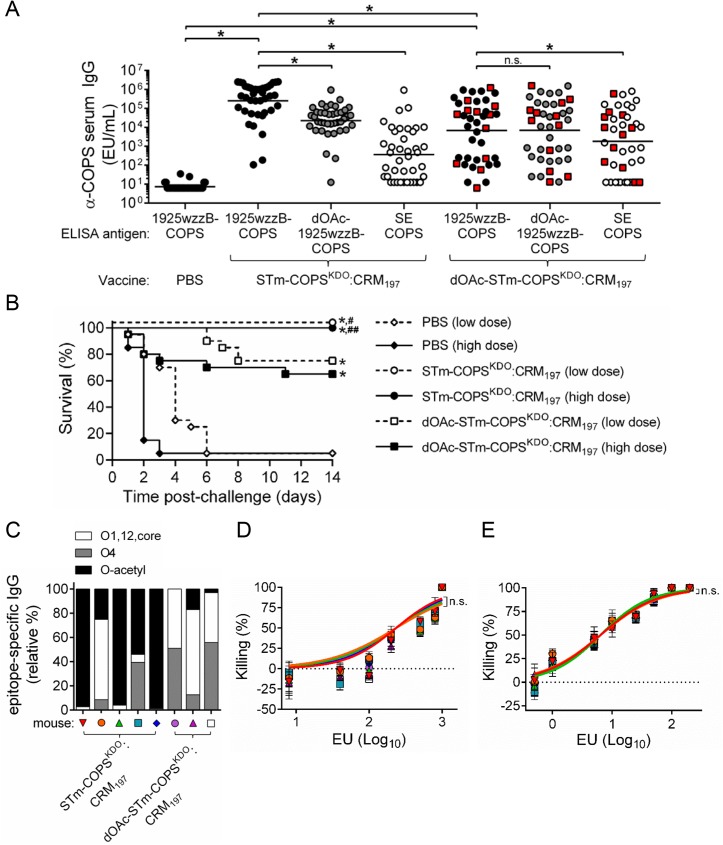
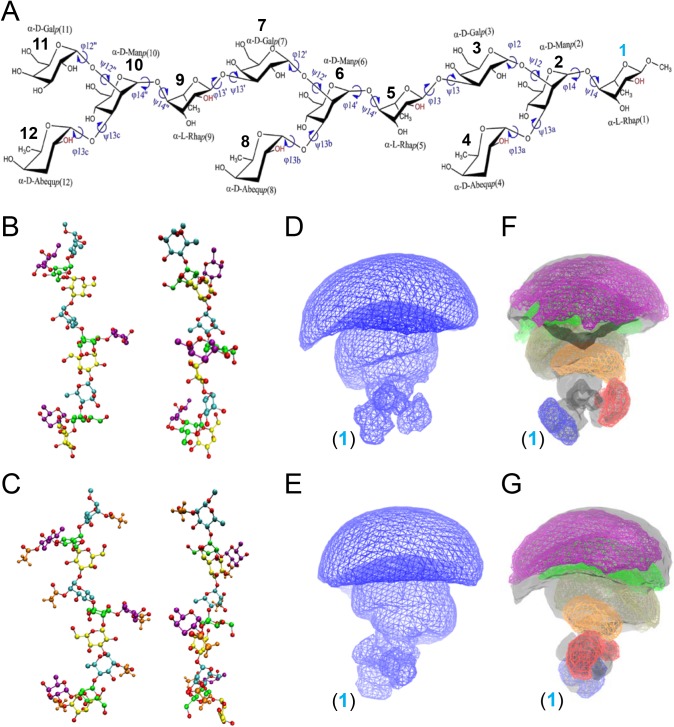
Invasive infections associated with non-typhoidal Salmonella (NTS) serovars Enteritidis (SE), Typhimurium (STm) and monophasic variant 1,4,[5],12:i:- are a major health problem in infants and young children in sub-Saharan Africa, and currently, there are no approved human NTS vaccines. NTS O-polysaccharides and flagellin proteins are protective antigens in animal models of invasive NTS infection. Conjugates of SE core and O-polysaccharide (COPS) chemically linked to SE flagellin have enhanced the anti-COPS immune response and protected mice against fatal challenge with a Malian SE blood isolate. We report herein the development of a STm glycoconjugate vaccine comprised of STm COPS conjugated to the homologous serovar phase 1 flagellin protein (FliC) with assessment of the role of COPS O-acetyls for functional immunity. Sun-type COPS conjugates linked through the polysaccharide reducing end to FliC were more immunogenic and protective in mice challenged with a Malian STm blood isolate than multipoint lattice conjugates (>95% vaccine efficacy [VE] versus 30-43% VE). Immunization with de-O-acetylated STm-COPS conjugated to CRM197 provided significant but reduced protection against STm challenge compared to mice immunized with native STm-COPS:CRM197 (63-74% VE versus 100% VE). Although OPS O-acetyls were highly immunogenic, post-vaccination sera that contained various O-acetyl epitope-specific antibody profiles displayed similar in vitro bactericidal activity when equivalent titers of anti-COPS IgG were assayed. In-silico molecular modeling further indicated that STm OPS forms a single dominant conformation, irrespective of O-acetylation, in which O-acetyls extend outward and are highly solvent exposed. These preclinical results establish important quality attributes for an STm vaccine that could be co-formulated with an SE-COPS:FliC glycoconjugate as a bivalent NTS vaccine for use in sub-Saharan Africa.
Conflict of interest statement
RS, SMT, JEG and MML are inventors on a patent describing flagellin based
Figures
References
-
- Berkley JA, Lowe BS, Mwangi I, Williams T, Bauni E, Mwarumba S, et al. Bacteremia among children admitted to a rural hospital in Kenya. N Engl J Med. 2005;352(1):39–47. Epub 2005/01/07. doi: 10.1056/NEJMoa040275 - DOI - PubMed
-
- Tapia MD, Tennant SM, Bornstein K, Onwuchekwa U, Tamboura B, Maiga A, et al. Invasive nontyphoidal Salmonella infections among children in Mali, 2002–2014: microbiological and epidemiologic features guide vaccine development. Clin Infect Dis. 2015;61 Suppl 4:S332–8. PubMed Central PMCID: PMCPMC4596934. - PMC - PubMed
-
- Okoro CK, Kingsley RA, Connor TR, Harris SR, Parry CM, Al-Mashhadani MN, et al. Intracontinental spread of human invasive Salmonella Typhimurium pathovariants in sub-Saharan Africa. Nat Genet. 2012;44(11):1215–21. PubMed Central PMCID: PMCPMC3491877. doi: 10.1038/ng.2423 - DOI - PMC - PubMed
-
- Kingsley RA, Msefula CL, Thomson NR, Kariuki S, Holt KE, Gordon MA, et al. Epidemic multiple drug resistant Salmonella Typhimurium causing invasive disease in sub-Saharan Africa have a distinct genotype. Genome Res. 2009;19(12):2279–87. PubMed Central PMCID: PMC2792184. doi: 10.1101/gr.091017.109 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
