

The bHLH transcription factor SPATULA enables cytokinin signaling, and both activate auxin biosynthesis and transport genes at the medial domain of the gynoecium
- PMID: 28388635
- PMCID: PMC5400277
- DOI: 10.1371/journal.pgen.1006726
The bHLH transcription factor SPATULA enables cytokinin signaling, and both activate auxin biosynthesis and transport genes at the medial domain of the gynoecium
Abstract
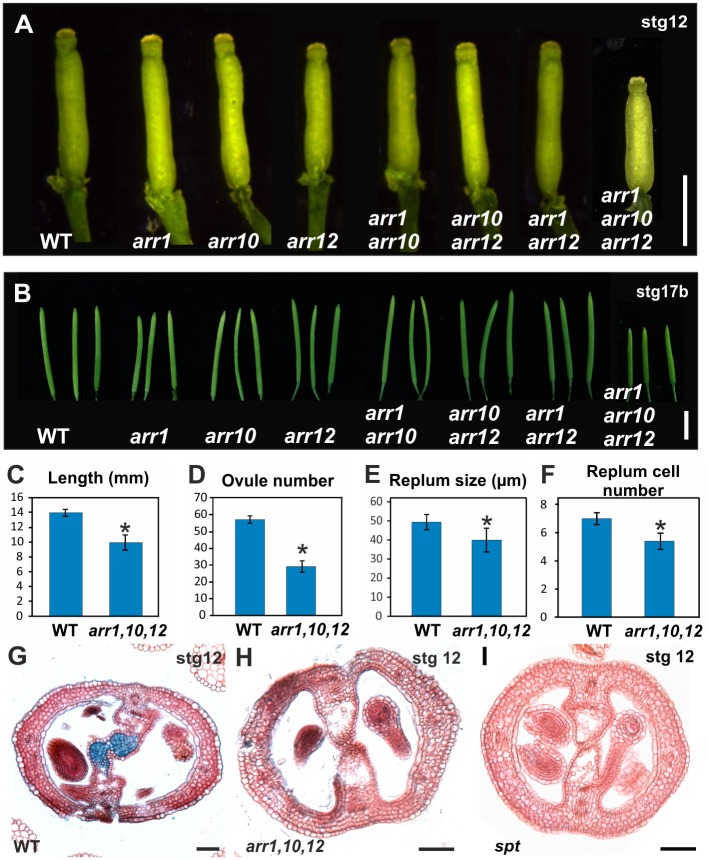
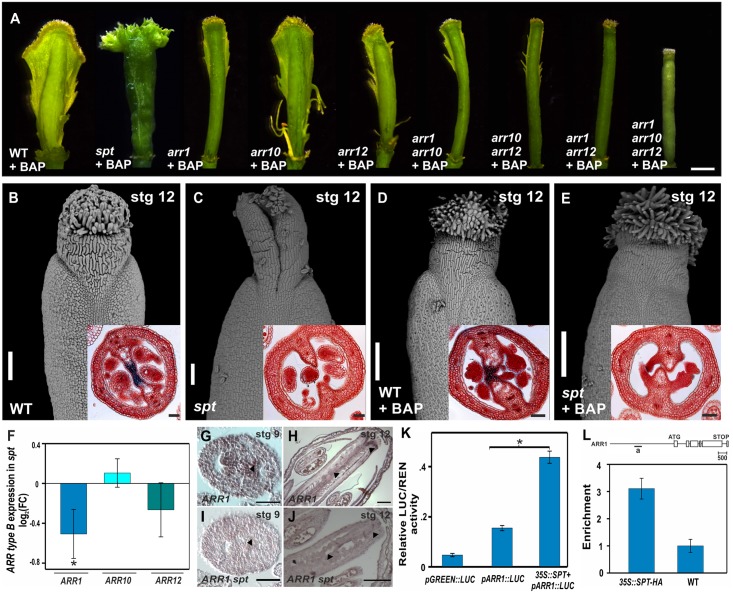
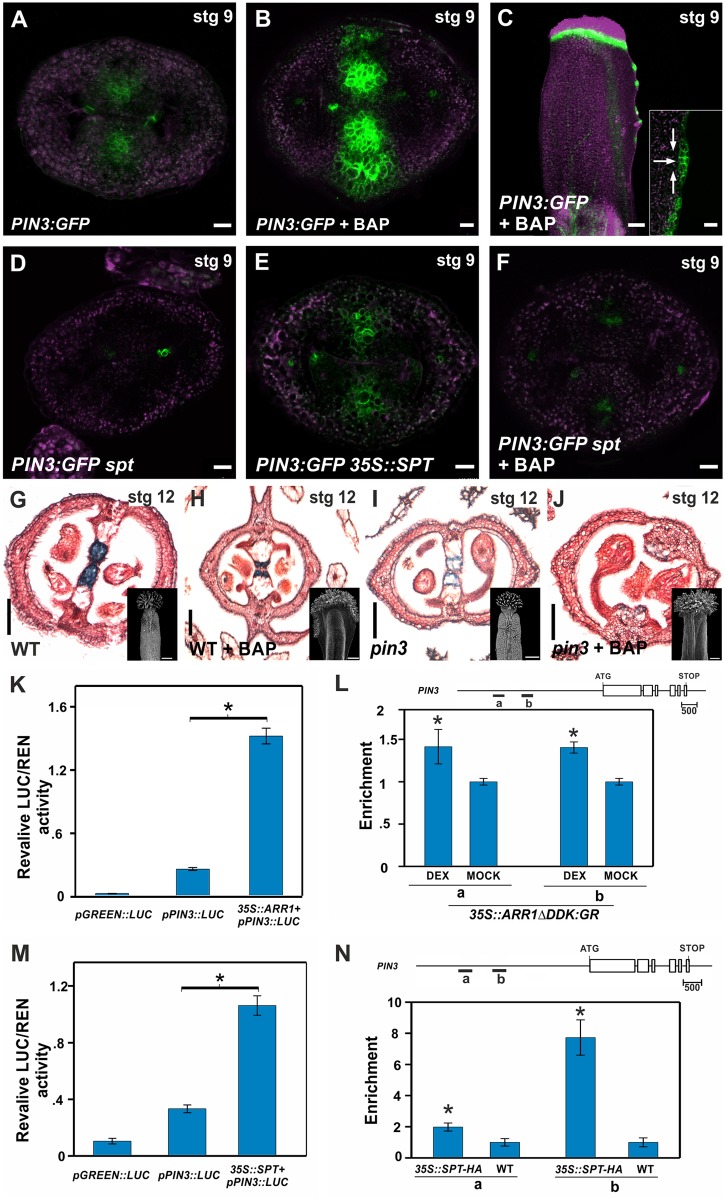
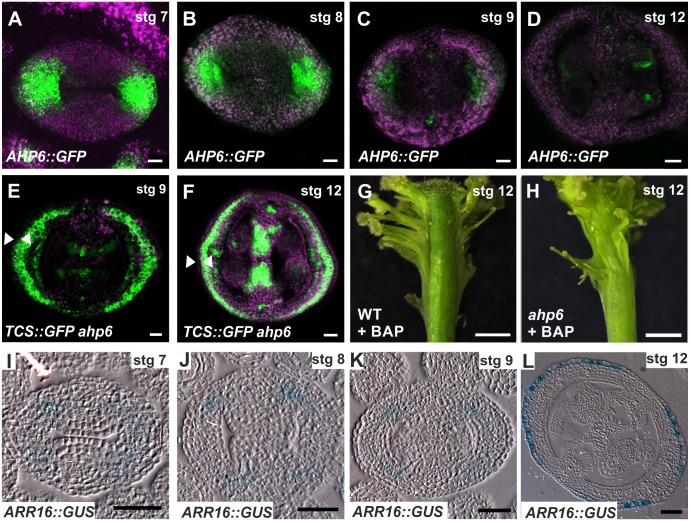
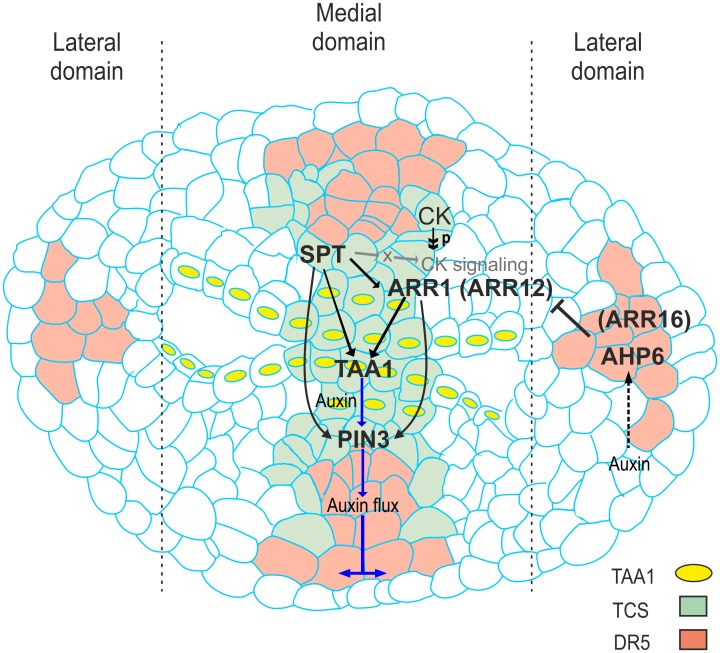
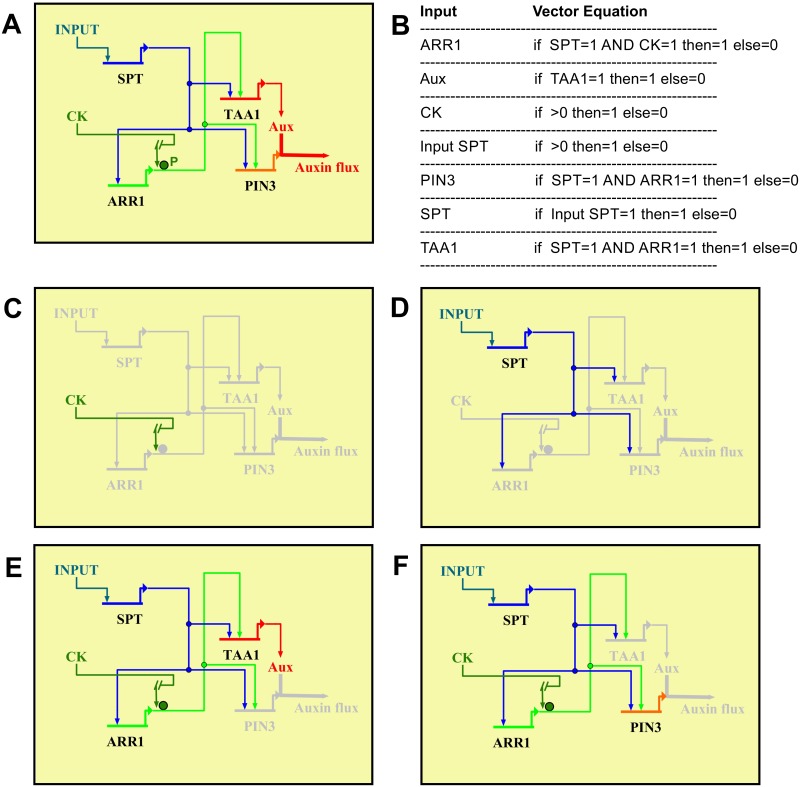
Fruits and seeds are the major food source on earth. Both derive from the gynoecium and, therefore, it is crucial to understand the mechanisms that guide the development of this organ of angiosperm species. In Arabidopsis, the gynoecium is composed of two congenitally fused carpels, where two domains: medial and lateral, can be distinguished. The medial domain includes the carpel margin meristem (CMM) that is key for the production of the internal tissues involved in fertilization, such as septum, ovules, and transmitting tract. Interestingly, the medial domain shows a high cytokinin signaling output, in contrast to the lateral domain, where it is hardly detected. While it is known that cytokinin provides meristematic properties, understanding on the mechanisms that underlie the cytokinin signaling pattern in the young gynoecium is lacking. Moreover, in other tissues, the cytokinin pathway is often connected to the auxin pathway, but we also lack knowledge about these connections in the young gynoecium. Our results reveal that cytokinin signaling, that can provide meristematic properties required for CMM activity and growth, is enabled by the transcription factor SPATULA (SPT) in the medial domain. Meanwhile, cytokinin signaling is confined to the medial domain by the cytokinin response repressor ARABIDOPSIS HISTIDINE PHOSPHOTRANSFERASE 6 (AHP6), and perhaps by ARR16 (a type-A ARR) as well, both present in the lateral domains (presumptive valves) of the developing gynoecia. Moreover, SPT and cytokinin, probably together, promote the expression of the auxin biosynthetic gene TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS 1 (TAA1) and the gene encoding the auxin efflux transporter PIN-FORMED 3 (PIN3), likely creating auxin drainage important for gynoecium growth. This study provides novel insights in the spatiotemporal determination of the cytokinin signaling pattern and its connection to the auxin pathway in the young gynoecium.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


Similar articles
-
Cytokinin-Auxin Crosstalk in the Gynoecial Primordium Ensures Correct Domain Patterning.Plant Physiol. 2017 Nov;175(3):1144-1157. doi: 10.1104/pp.17.00805. Epub 2017 Sep 11. Plant Physiol. 2017. PMID: 28894023 Free PMC article.
-
The role of D3-type cyclins is related to cytokinin and the bHLH transcription factor SPATULA in Arabidopsis gynoecium development.Planta. 2024 Jul 9;260(2):48. doi: 10.1007/s00425-024-04481-4. Planta. 2024. PMID: 38980389 Free PMC article.
-
Synergistic relationship between auxin and cytokinin in the ovary and the participation of the transcription factor SPATULA.Plant Signal Behav. 2017 Oct 3;12(10):e1376158. doi: 10.1080/15592324.2017.1376158. Epub 2017 Sep 7. Plant Signal Behav. 2017. PMID: 28880725 Free PMC article.
-
Auxin and cytokinin act during gynoecial patterning and the development of ovules from the meristematic medial domain.Wiley Interdiscip Rev Dev Biol. 2015 Nov-Dec;4(6):555-71. doi: 10.1002/wdev.193. Epub 2015 May 7. Wiley Interdiscip Rev Dev Biol. 2015. PMID: 25951007 Review.
-
Inside the gynoecium: at the carpel margin.Trends Plant Sci. 2013 Nov;18(11):644-55. doi: 10.1016/j.tplants.2013.08.002. Epub 2013 Sep 2. Trends Plant Sci. 2013. PMID: 24008116 Review.
Cited by
-
ADA2b and GCN5 Affect Cytokinin Signaling by Modulating Histone Acetylation and Gene Expression during Root Growth of Arabidopsis thaliana.Plants (Basel). 2022 May 18;11(10):1335. doi: 10.3390/plants11101335. Plants (Basel). 2022. PMID: 35631760 Free PMC article.
-
Biosynthesis, Metabolism and Function of Auxin, Salicylic Acid and Melatonin in Climacteric and Non-climacteric Fruits.Front Plant Sci. 2019 Feb 18;10:136. doi: 10.3389/fpls.2019.00136. eCollection 2019. Front Plant Sci. 2019. PMID: 30833953 Free PMC article. Review.
-
DNA methylation is involved in sexual differentiation and sex chromosome evolution in the dioecious plant garden asparagus.Hortic Res. 2021 Sep 1;8(1):198. doi: 10.1038/s41438-021-00633-9. Hortic Res. 2021. PMID: 34465747 Free PMC article.
-
Precise Regulation of the TAA1/TAR-YUCCA Auxin Biosynthesis Pathway in Plants.Int J Mol Sci. 2023 May 10;24(10):8514. doi: 10.3390/ijms24108514. Int J Mol Sci. 2023. PMID: 37239863 Free PMC article. Review.
-
EXPANSIN15 is involved in flower and fruit development in Arabidopsis.Plant Reprod. 2024 Jun;37(2):259-270. doi: 10.1007/s00497-023-00493-4. Epub 2024 Jan 29. Plant Reprod. 2024. PMID: 38285171 Free PMC article.
References
-
- Bowman JL, Baum SF, Eshed Y, Putterill J, Alvarez J (1999) Molecular genetics of gynoecium development in Arabidopsis. Curr Top Dev Biol 45: 155–205. - PubMed
-
- Chávez Montes RA, Herrera-Ubaldo H, Serwatowska J, de Folter S (2015) Towards a comprehensive and dynamic gynoecium gene regulatory network. Current Plant Biology 3–4: 3–12.
-
- Sehra B, Franks RG (2015) Auxin and cytokinin act during gynoecial patterning and the development of ovules from the meristematic medial domain. Wiley Interdiscip Rev Dev Biol. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
