

Theoretical and Biological Evaluation of the Link between Low Exercise Capacity and Disease Risk
- PMID: 28389512
- PMCID: PMC5749140
- DOI: 10.1101/cshperspect.a029868
Theoretical and Biological Evaluation of the Link between Low Exercise Capacity and Disease Risk
Abstract
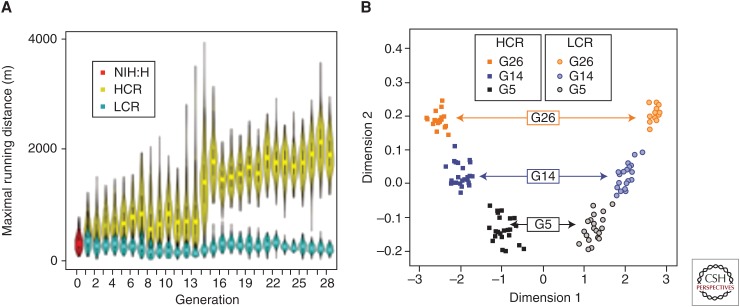
Large-scale epidemiological studies show that low exercise capacity is the highest risk factor for all-cause morbidity and mortality relative to other conditions including diabetes, hypertension, and obesity. This led us to formulate the energy transfer hypothesis (ETH): Variation in capacity for energy transfer is the central mechanistic determinant of the divide between disease and health. As a test of this hypothesis, we predicted that two-way selective breeding of genetically heterogeneous rats for low and high intrinsic treadmill running capacity (a surrogate for energy transfer) would also produce rats that differ for disease risks. The lines are termed low-capacity runners (LCRs) and high-capacity runners (HCRs) and, after 36 generations of selection, they differ by more than eightfold in running capacity. Consistent with the ETH, the LCRs score high for developing disease risks, including metabolic syndrome, neurodegeneration, cognitive impairment, fatty liver disease, susceptibility to cancer, and reduced longevity. The HCRs are resistant to the development of these disease risks. Here we synthesize ideas on nonequilibrium thermodynamics and evolution from Ilya Prigogine, Hans Krebs, and Peter Mitchell to formulate theoretic explanations for the ETH. First, at every moment in time, the atoms and molecules of organisms are reorganizing to pursue avenues for energy transfer. Second, this continuous organization is navigating in a constantly changing environment such that "strategies" are perpetually in flux and do not leave a simple footprint (evolution). Third, as a consequence, human populations demonstrate a wide variation in capacity for energy transfer that mirrors mechanistically the divide between disease and health.
Copyright © 2018 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Atamian HS, Creux NM, Brown EA, Garner AG, Blackman BK, Harmer SL. 2016. Circadian regulation of sunflower heliotropism, floral orientation, and pollinator visits. Science 353: 587–590. - PubMed
-
- Baldwin JE, Krebs H. 1981. The evolution of metabolic cycles. Nature 291: 381–382. - PubMed
-
- Bonow RO, Borer JS, Rosing DR, Henry WL, Pearlman AS, McIntosh CL, Morrow AG, Epstein SE. 1980. Preoperative exercise capacity in symptomatic patients with aortic regurgitation as a predictor of postoperative left ventricular function and long-term prognosis. Circulation 62: 1280–1290. - PubMed
-
- Bortz WM II. 2015. Metabolic field (Schrodinger); an explanatory platform for biology: Based on lecture at Trinity College, Dublin, Ireland, July 18, 2012. Med Hypotheses 85: 894–897. - PubMed
-
- Bouchard C, Rankinen T, Chagnon YC, Rice T, Perusse L, Gagnon J, Borecki I, An P, Leon AS, Skinner JS, et al. 2000. Genomic scan for maximal oxygen uptake and its response to training in the HERITAGE family study. J Appl Physiol 88: 551–559. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials