

The dual developmental origin of spinal cerebrospinal fluid-contacting neurons gives rise to distinct functional subtypes
- PMID: 28389647
- PMCID: PMC5428266
- DOI: 10.1038/s41598-017-00350-1
The dual developmental origin of spinal cerebrospinal fluid-contacting neurons gives rise to distinct functional subtypes
Abstract
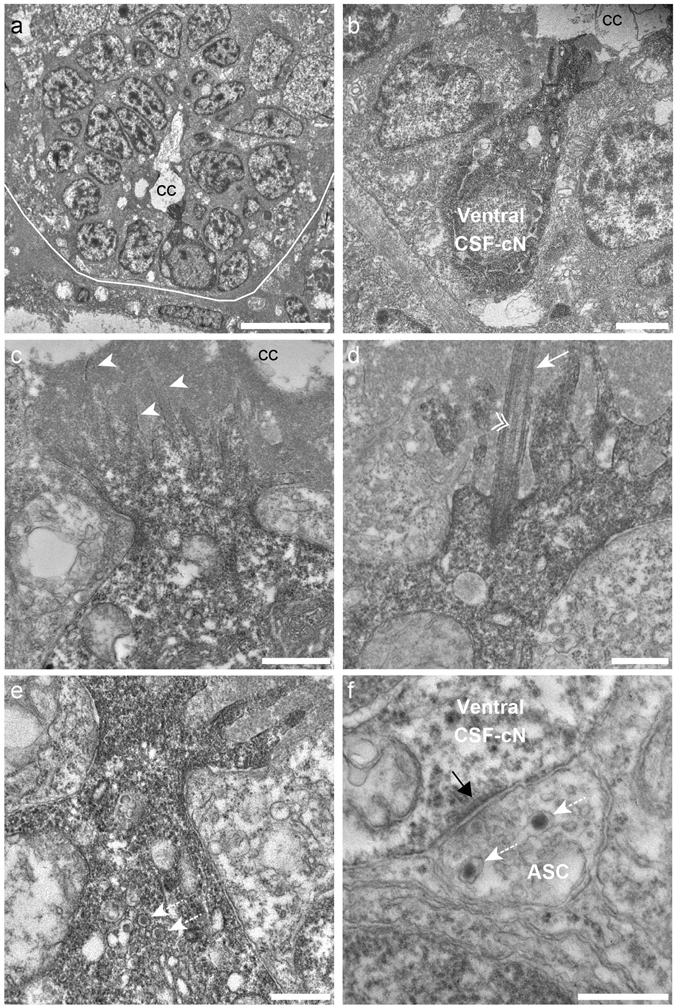
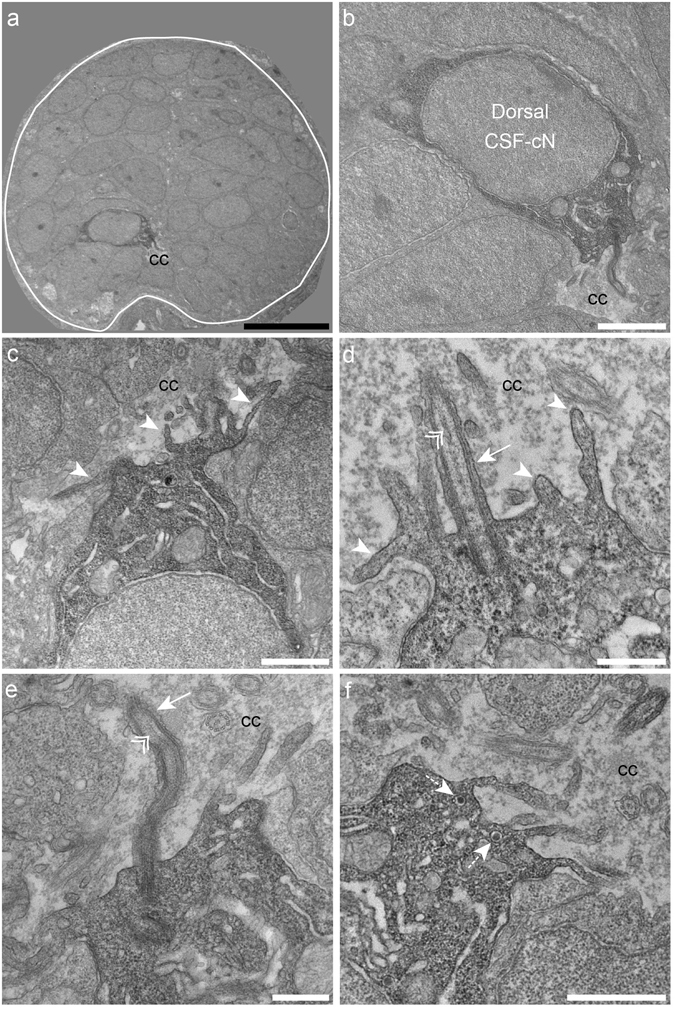
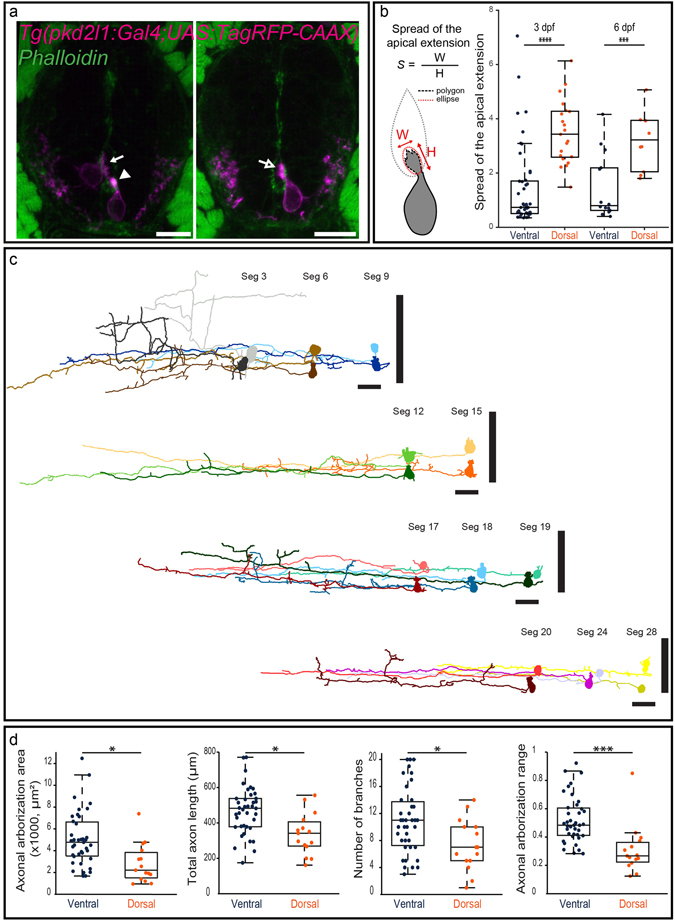
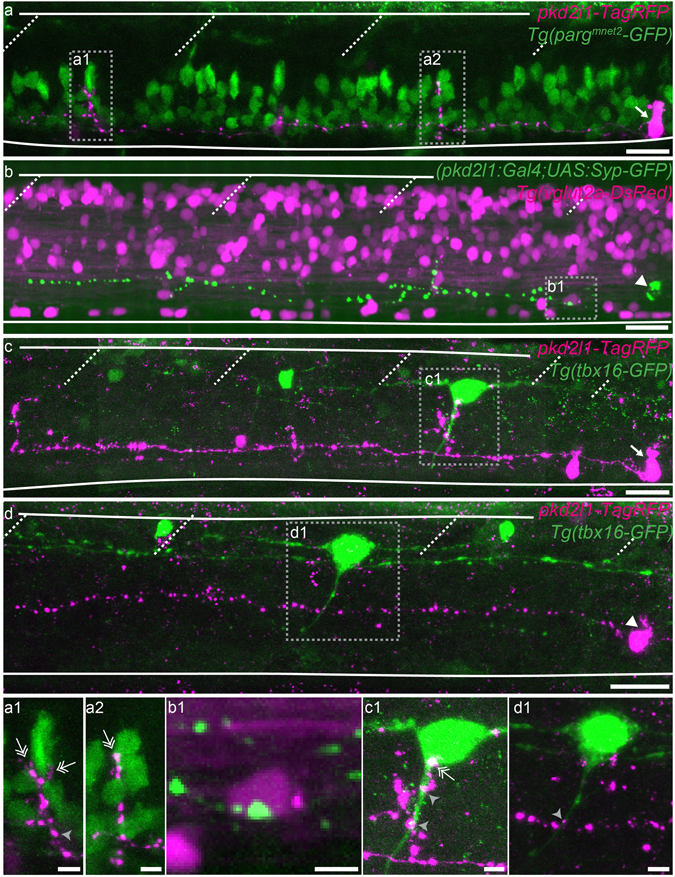
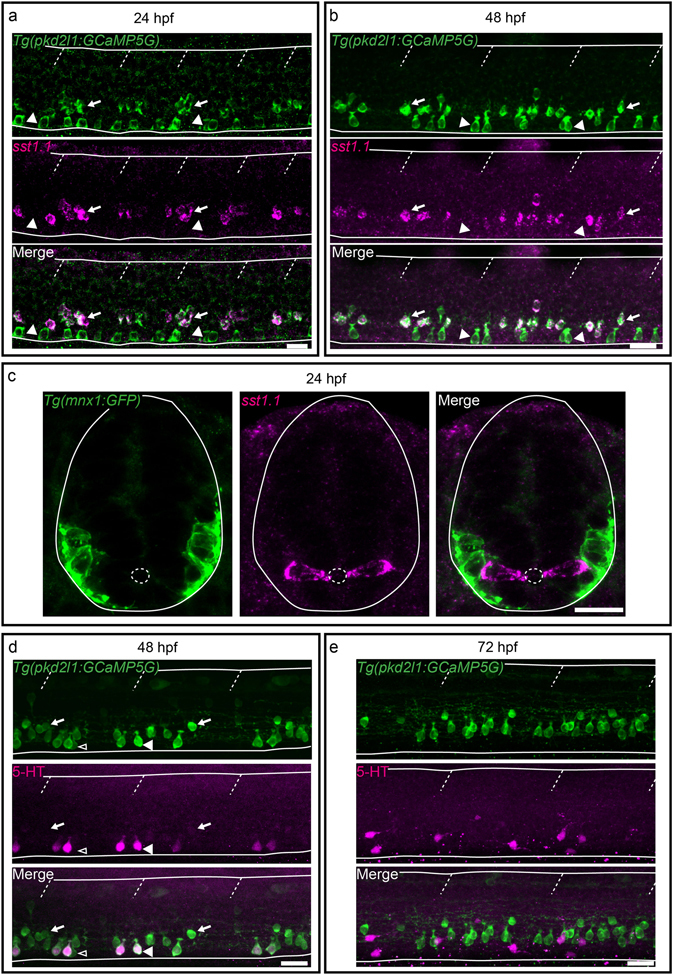
Chemical and mechanical cues from the cerebrospinal fluid (CSF) can affect the development and function of the central nervous system (CNS). How such cues are detected and relayed to the CNS remains elusive. Cerebrospinal fluid-contacting neurons (CSF-cNs) situated at the interface between the CSF and the CNS are ideally located to convey such information to local networks. In the spinal cord, these GABAergic neurons expressing the PKD2L1 channel extend an apical extension into the CSF and an ascending axon in the spinal cord. In zebrafish and mouse spinal CSF-cNs originate from two distinct progenitor domains characterized by distinct cascades of transcription factors. Here we ask whether these neurons with different developmental origins differentiate into cells types with different functional properties. We show in zebrafish larva that the expression of specific markers, the morphology of the apical extension and axonal projections, as well as the neuronal targets contacted by CSF-cN axons, distinguish the two CSF-cN subtypes. Altogether our study demonstrates that the developmental origins of spinal CSF-cNs give rise to two distinct functional populations of sensory neurons. This work opens novel avenues to understand how these subtypes may carry distinct functions related to development of the spinal cord, locomotion and posture.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
