

Involvement of salicylic acid, ethylene and jasmonic acid signalling pathways in the susceptibility of tomato to Fusarium oxysporum
- PMID: 28390170
- PMCID: PMC6638294
- DOI: 10.1111/mpp.12559
Involvement of salicylic acid, ethylene and jasmonic acid signalling pathways in the susceptibility of tomato to Fusarium oxysporum
Abstract
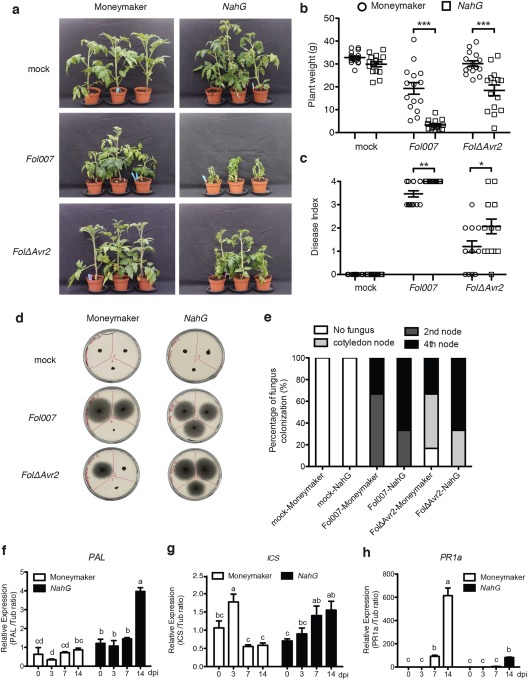
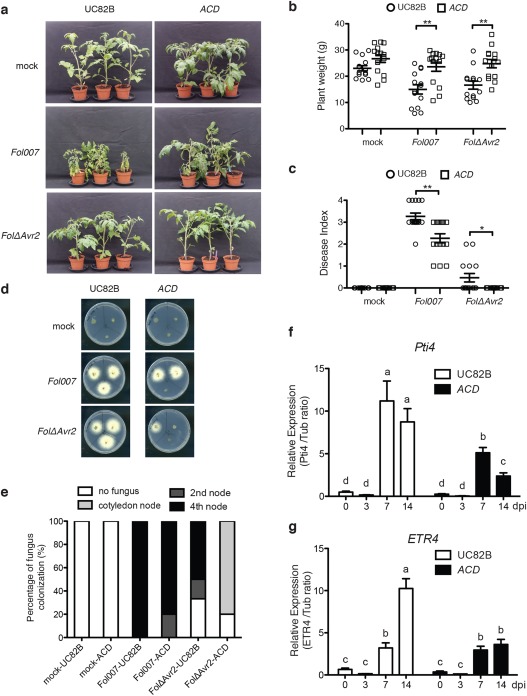
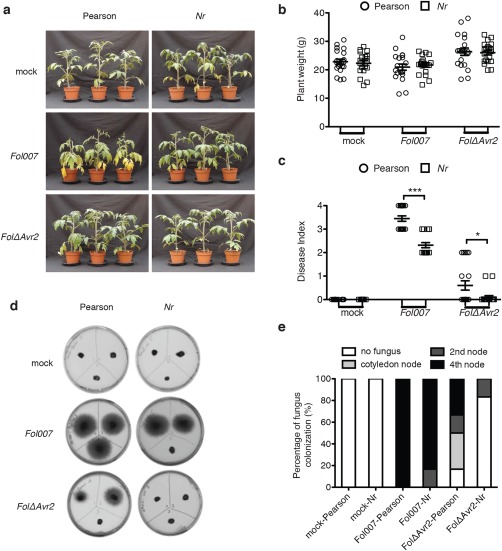
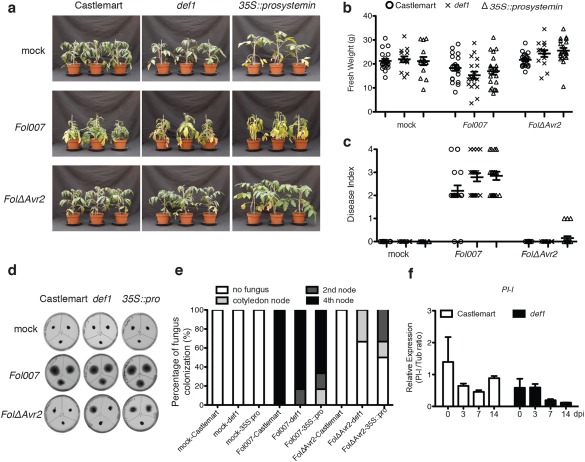
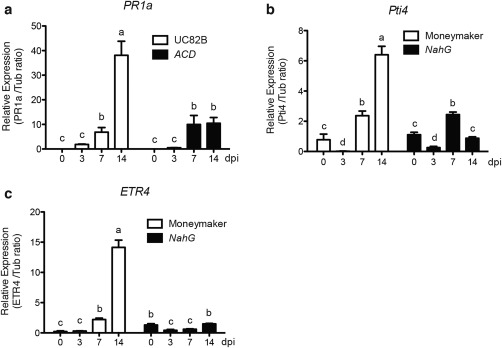
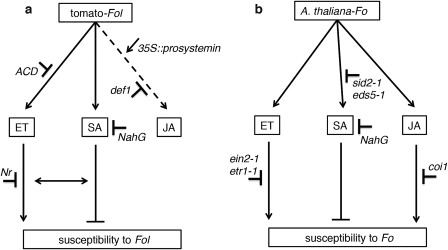
Phytohormones, such as salicylic acid (SA), ethylene (ET) and jasmonic acid (JA), play key roles in plant defence following pathogen attack. The involvement of these hormones in susceptibility following Fusarium oxysporum (Fo) infection has mostly been studied in Arabidopsis thaliana. However, Fo causes vascular wilt disease in a broad range of crops, including tomato (Solanum lycopersicum). Surprisingly little is known about the involvement of these phytohormones in the susceptibility of tomato towards Fo f. sp. lycopersici (Fol). Here, we investigate their involvement by the analysis of the expression of ET, JA and SA marker genes following Fol infection, and by bioassays of tomato mutants affected in either hormone production or perception. Fol inoculation triggered the expression of SA and ET marker genes, showing the activation of these pathways. NahG tomato, in which SA is degraded, became hypersusceptible to Fol infection and showed stronger disease symptoms than wild-type. In contrast, ACD and Never ripe (Nr) mutants, in which ET biosynthesis and perception, respectively, are impaired, showed decreased disease symptoms and reduced fungal colonization on infection. The susceptibility of the def1 tomato mutant, and a prosystemin over-expressing line, in which JA signalling is compromised or constitutively activated, respectively, was unaltered. Our results show that SA is a negative and ET a positive regulator of Fol susceptibility. The SA and ET signalling pathways appear to act synergistically, as an intact ET pathway is required for the induction of an SA marker gene, and vice versa.
Keywords: ET; Fusarium oxysporum; JA; SA; susceptibility; tomato.
© 2017 THE AUTHORS. MOLECULAR PLANT PATHOLOGY PUBLISHED BY BRITISH SOCIETY FOR PLANT PATHOLOGY AND JOHN WILEY & SONS LTD.
Figures
References
-
- Agrios, G.N. (2005) Plant Pathology. St. Louis, MO: Academic Press.
-
- Anderson, J.P. , Badruzsaufari, E. , Schenk, P.M. , Manners, J.M. , Desmond, O.J. , Ehlert, C. , Maclean, D.J. , Ebert, P.R. and Kazan, K. (2004) Antagonistic interaction between abscisic acid and jasmonate–ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell, 16, 3460–3479. - PMC - PubMed
-
- Bari, R. and Jones, J.D.G. (2009) Role of plant hormones in plant defence responses. Plant Mol. Biol. 69, 473–488. - PubMed
-
- Berrocal‐Lobo, M. and Molina, A. (2004) Ethylene response factor 1 mediates Arabidopsis resistance to the soilborne fungus Fusarium oxysporum . Mol. Plant–Microbe Interact. 17, 763–770. - PubMed
-
- Berrocal‐Lobo, M. and Molina, A. (2008) Arabidopsis defense response against Fusarium oxysporum . Trends Plant Sci. 13, 145–150. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
