

Evolutionary Thrift: Mycobacteria Repurpose Plasmid Diversity during Adaptation of Type VII Secretion Systems
- PMID: 28391322
- PMCID: PMC5381665
- DOI: 10.1093/gbe/evx001
Evolutionary Thrift: Mycobacteria Repurpose Plasmid Diversity during Adaptation of Type VII Secretion Systems
Abstract
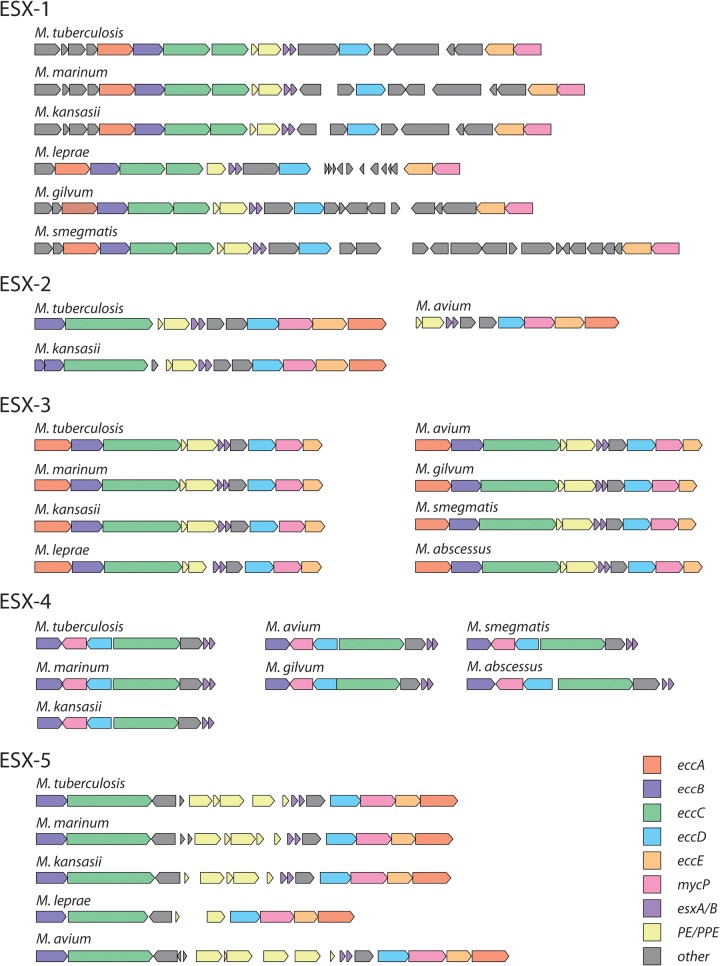
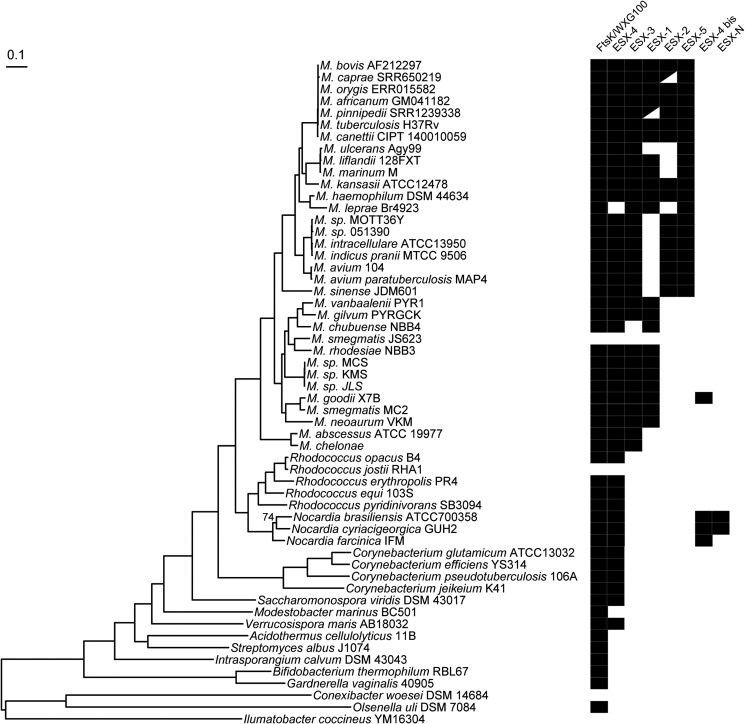
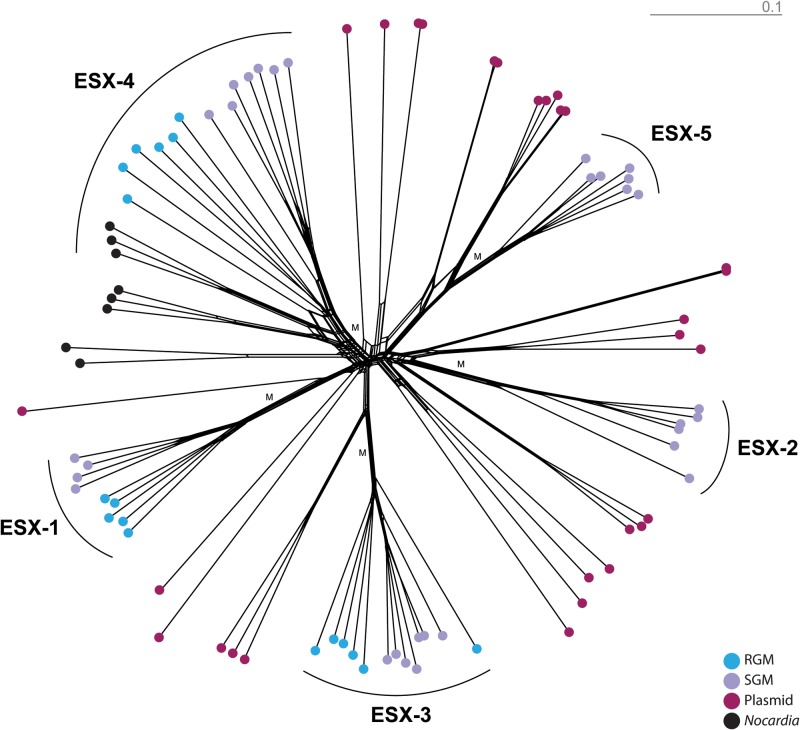
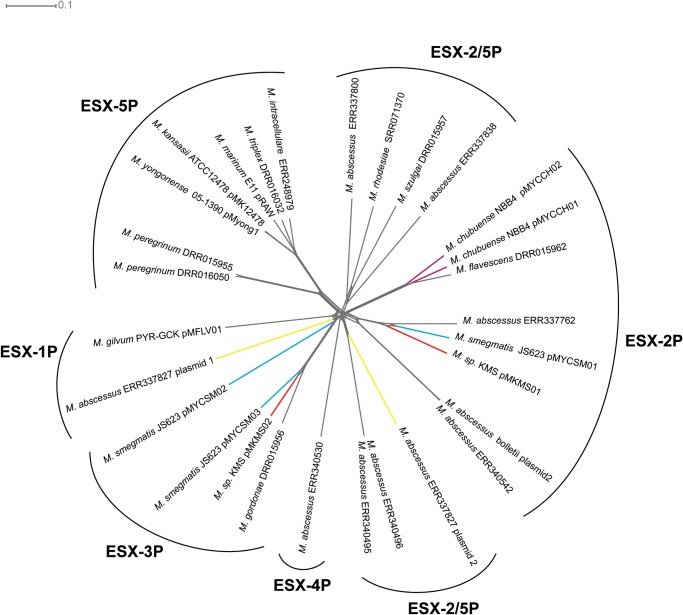
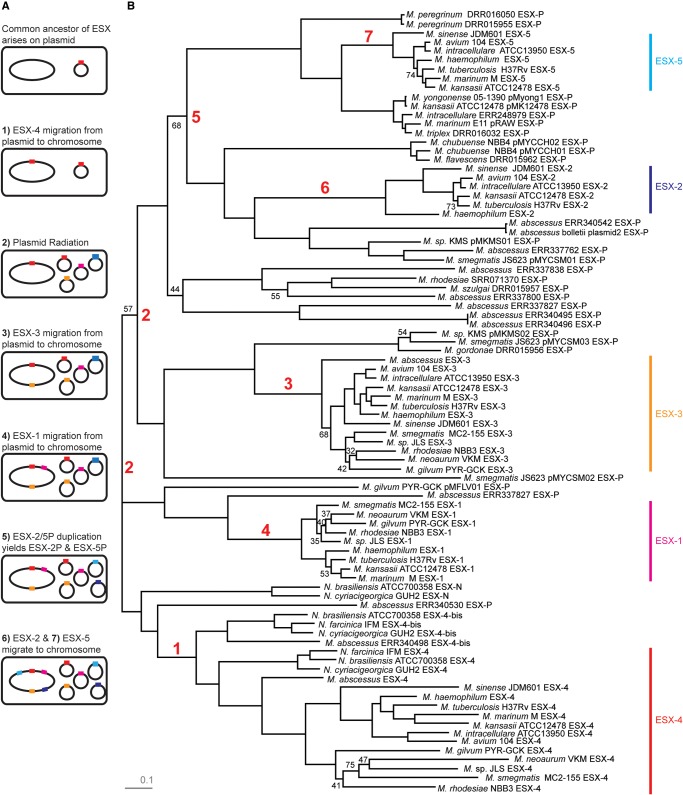
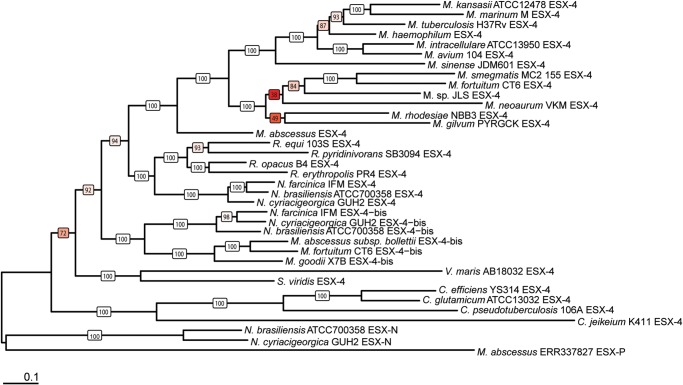
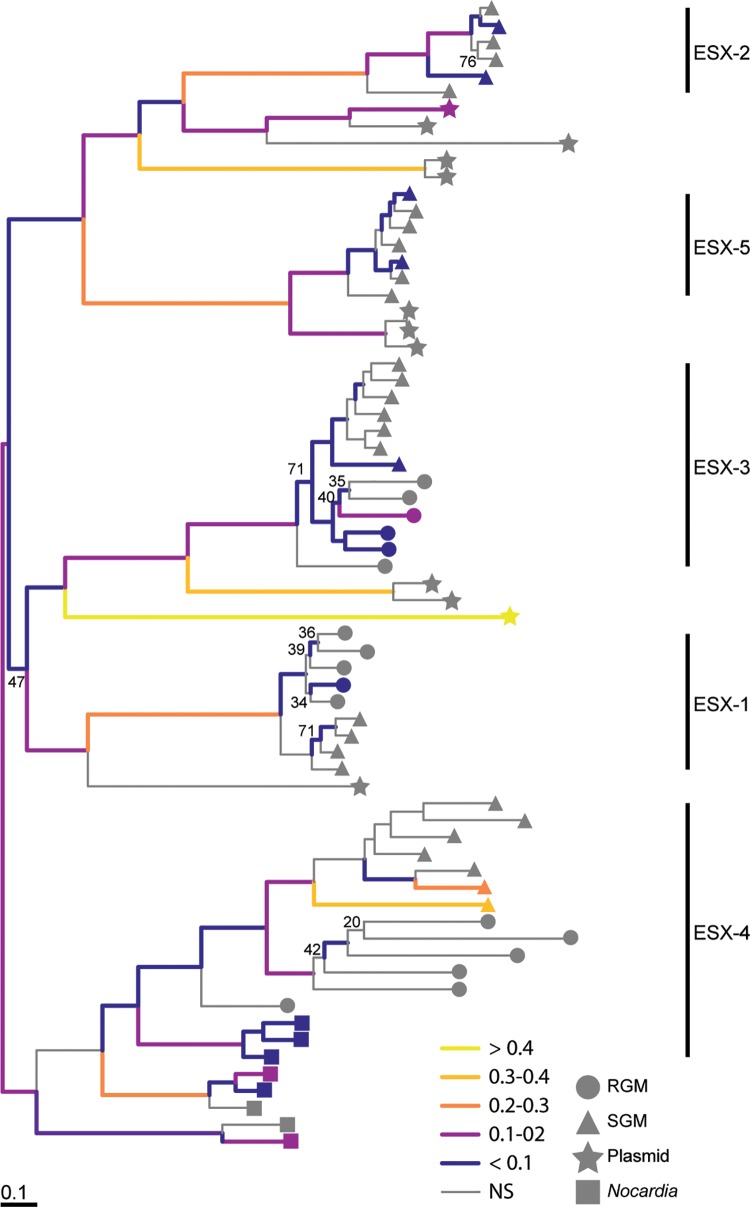
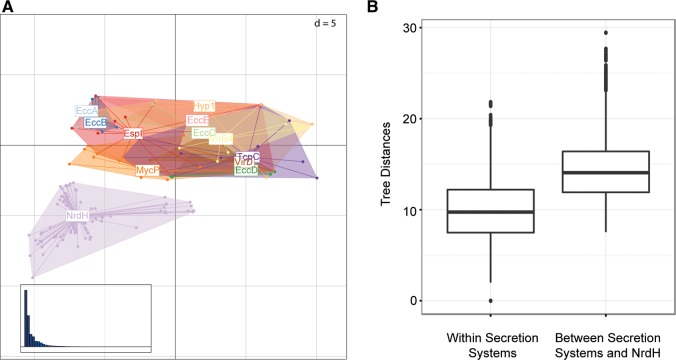
Mycobacteria have a distinct secretion system, termed type VII (T7SS), which is encoded by paralogous chromosomal loci (ESX) and associated with pathogenesis, conjugation, and metal homeostasis. Evolution of paralogous gene families is of interest because duplication is an important mechanism by which novel genes evolve, but there are potential conflicts between adaptive forces that stabilize duplications and those that enable evolution of new functions. Our objective was to delineate the adaptive forces underlying diversification of T7SS. Plasmid-borne ESX were described recently, and we found evidence that the initial duplication and divergence of ESX systems occurred on plasmids and was driven by selection for advantageous mutations. Plasmid conjugation has been linked to T7SS and type IV secretion systems (T4SS) in mycobacteria, and we discovered that T7SS and T4SS genes evolved in concert on the plasmids. We hypothesize that differentiation of plasmid ESX helps to prevent conjugation among cells harboring incompatible plasmids. Plasmid ESX appear to have been repurposed following migration to the chromosome, and there is evidence of positive selection driving further differentiation of chromosomal ESX. We hypothesize that ESX loci were initially stabilized on the chromosome by mediating their own transfer. These results emphasize the diverse adaptive paths underlying evolution of novelty, which in this case involved plasmid duplications, selection for advantageous mutations in the mobile and core genomes, migration of the loci between plasmids and chromosomes, and lateral transfer among chromosomes. We discuss further implications for the choice of model organism to study ESX functions in Mycobacterium tuberculosis.
Keywords: ESX; gene duplication; mycobacteria; plasmid; selection; type VII secretion system.
© The Author(s) 2017. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
Similar articles
-
Multiple genetic paths including massive gene amplification allow Mycobacterium tuberculosis to overcome loss of ESX-3 secretion system substrates.Proc Natl Acad Sci U S A. 2022 Feb 22;119(8):e2112608119. doi: 10.1073/pnas.2112608119. Proc Natl Acad Sci U S A. 2022. PMID: 35193958 Free PMC article.
-
The plasmid-mediated evolution of the mycobacterial ESX (Type VII) secretion systems.BMC Evol Biol. 2016 Mar 15;16:62. doi: 10.1186/s12862-016-0631-2. BMC Evol Biol. 2016. PMID: 26979252 Free PMC article.
-
Mycobacterial Pan-Genome Analysis Suggests Important Role of Plasmids in the Radiation of Type VII Secretion Systems.Genome Biol Evol. 2016 Jan 8;8(2):387-402. doi: 10.1093/gbe/evw001. Genome Biol Evol. 2016. PMID: 26748339 Free PMC article.
-
Modular Organization of the ESX-5 Secretion System in Mycobacterium tuberculosis.Front Cell Infect Microbiol. 2016 May 2;6:49. doi: 10.3389/fcimb.2016.00049. eCollection 2016. Front Cell Infect Microbiol. 2016. PMID: 27200304 Free PMC article. Review.
-
Targeting type VII/ESX secretion systems for development of novel antimycobacterial drugs.Curr Pharm Des. 2014;20(27):4346-56. doi: 10.2174/1381612819666131118170717. Curr Pharm Des. 2014. PMID: 24245757 Review.
Cited by
-
Mycolicibacterium fortuitum genomic epidemiology, resistome and virulome.Mem Inst Oswaldo Cruz. 2022 Jan 10;116:e210247. doi: 10.1590/0074-02760210247. eCollection 2022. Mem Inst Oswaldo Cruz. 2022. PMID: 35019071 Free PMC article.
-
Plasmid-mediated drug resistance in mycobacteria: the tip of the iceberg?J Clin Microbiol. 2023 Oct 24;61(10):e0062823. doi: 10.1128/jcm.00628-23. Epub 2023 Sep 19. J Clin Microbiol. 2023. PMID: 37724858 Free PMC article.
-
Blending genomes: distributive conjugal transfer in mycobacteria, a sexier form of HGT.Mol Microbiol. 2018 Jun;108(6):601-613. doi: 10.1111/mmi.13971. Epub 2018 May 11. Mol Microbiol. 2018. PMID: 29669186 Free PMC article. Review.
-
Frequent transmission of the Mycobacterium tuberculosis Beijing lineage and positive selection for the EsxW Beijing variant in Vietnam.Nat Genet. 2018 Jun;50(6):849-856. doi: 10.1038/s41588-018-0117-9. Epub 2018 May 21. Nat Genet. 2018. PMID: 29785015 Free PMC article.
-
Multiple genetic paths including massive gene amplification allow Mycobacterium tuberculosis to overcome loss of ESX-3 secretion system substrates.Proc Natl Acad Sci U S A. 2022 Feb 22;119(8):e2112608119. doi: 10.1073/pnas.2112608119. Proc Natl Acad Sci U S A. 2022. PMID: 35193958 Free PMC article.
References
-
- Abdallah AM, et al. 2009. PPE and PE_PGRS proteins of Mycobacterium marinum are transported via the type VII secretion system ESX-5. Mol Microbiol. 73:329–340. doi: 10.1111/j.1365-2958.2009.06783.x. - PubMed
-
- Adler M, Anjum M, Berg OG, Andersson DI, Sandegren L. 2014. High fitness costs and instability of gene duplications reduce rates of evolution of new genes by duplication-divergence mechanisms. Mol Biol Evol. 31:1526–1535. - PubMed
-
- Andersson DI, Hughes D. 2009. Gene amplification and adaptive evolution in bacteria. Annu. Rev. Genet. 43:167–195. - PubMed
-
- Antipov D, et al. 2016. plasmidSPAdes: Assembling Plasmids from Whole Genome Sequencing Data. bioRxiv. 48942. doi: 10.1101/048942. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
