

The Evolution of Sex Chromosomes and Dosage Compensation in Plants
- PMID: 28391324
- PMCID: PMC5629387
- DOI: 10.1093/gbe/evw282
The Evolution of Sex Chromosomes and Dosage Compensation in Plants
Abstract
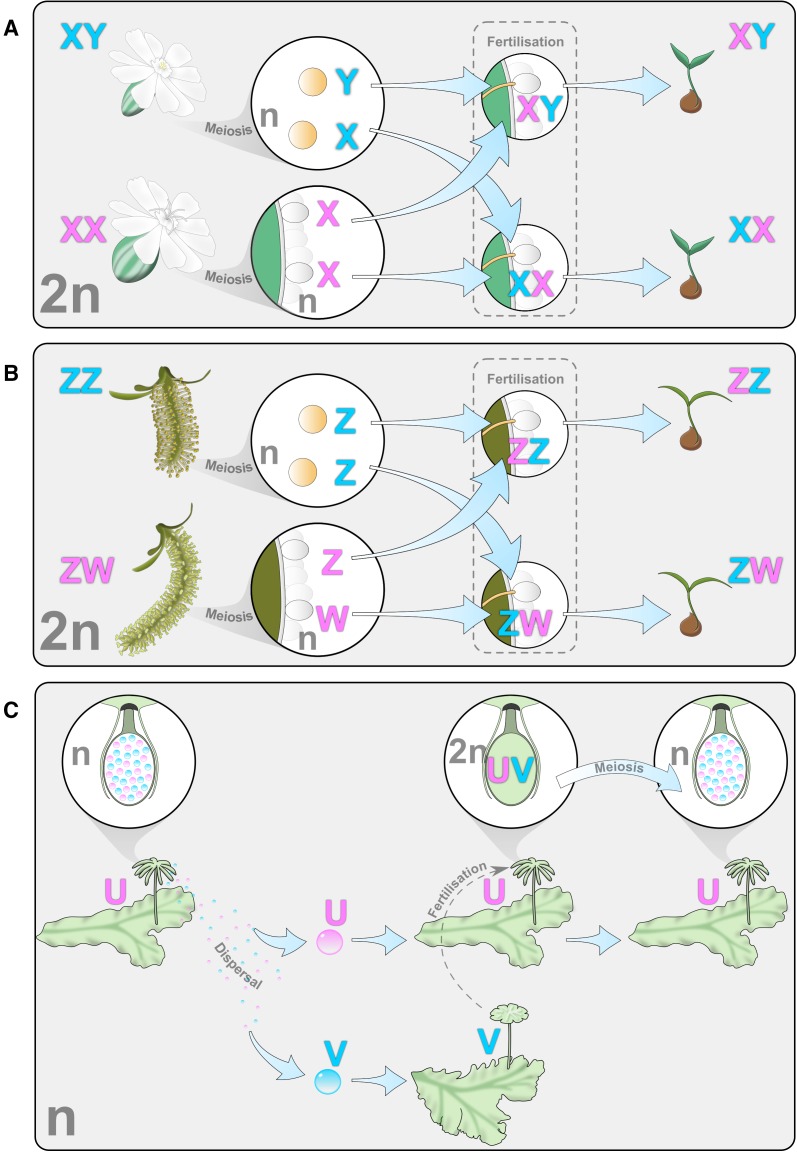
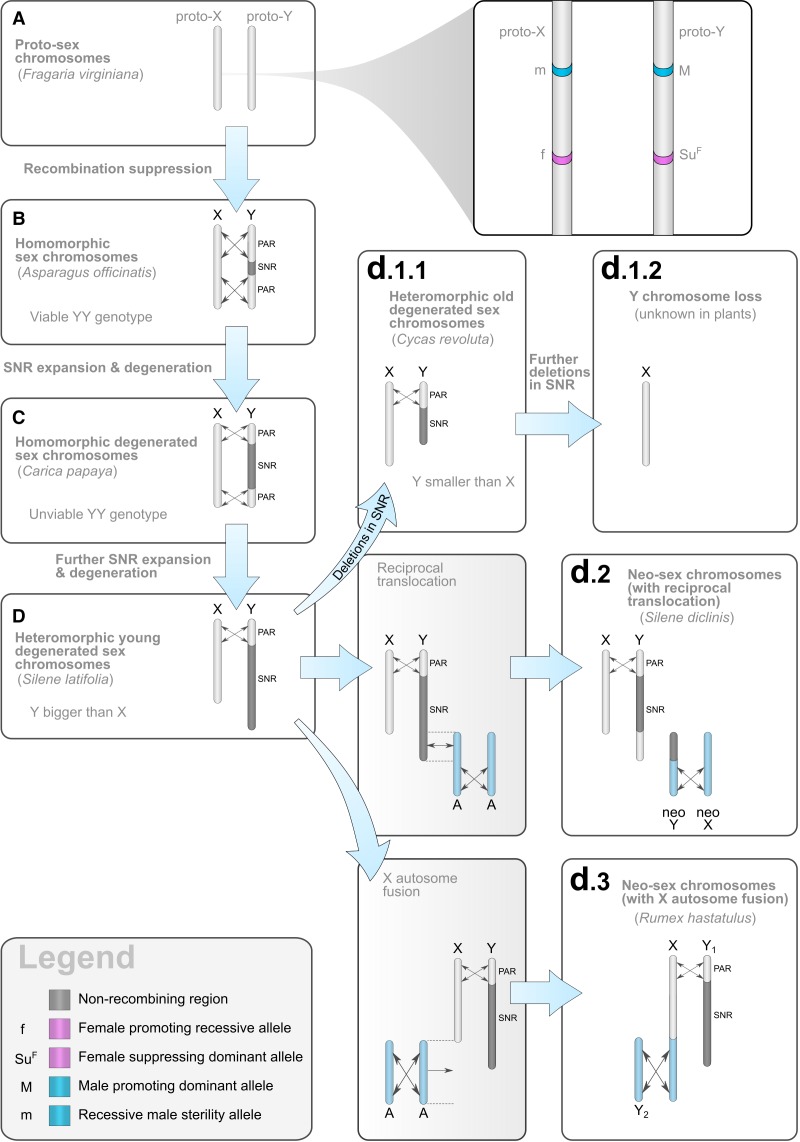
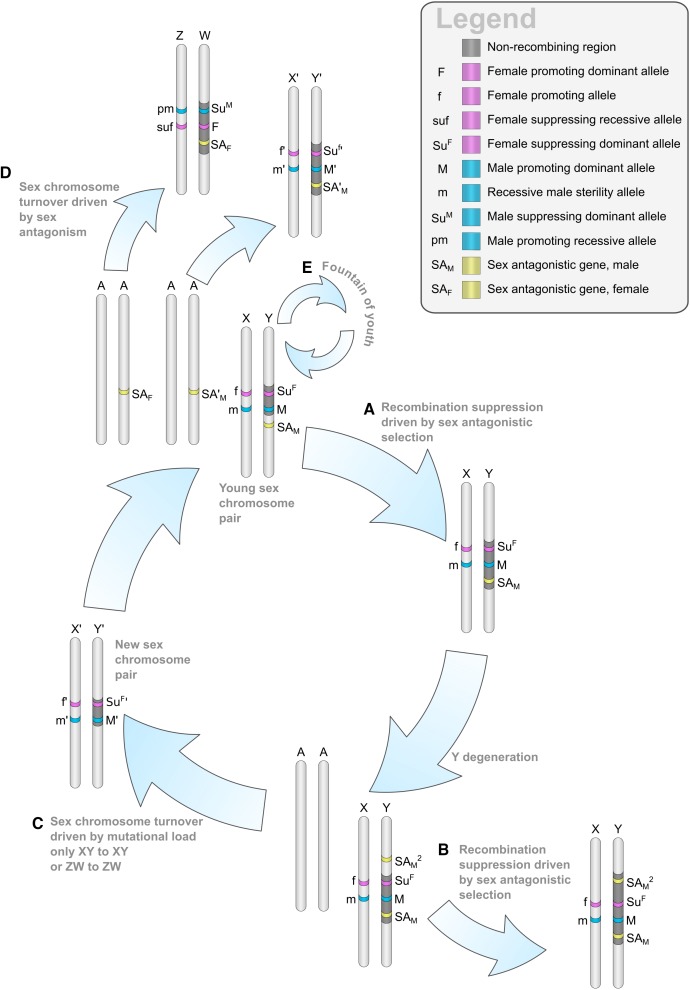
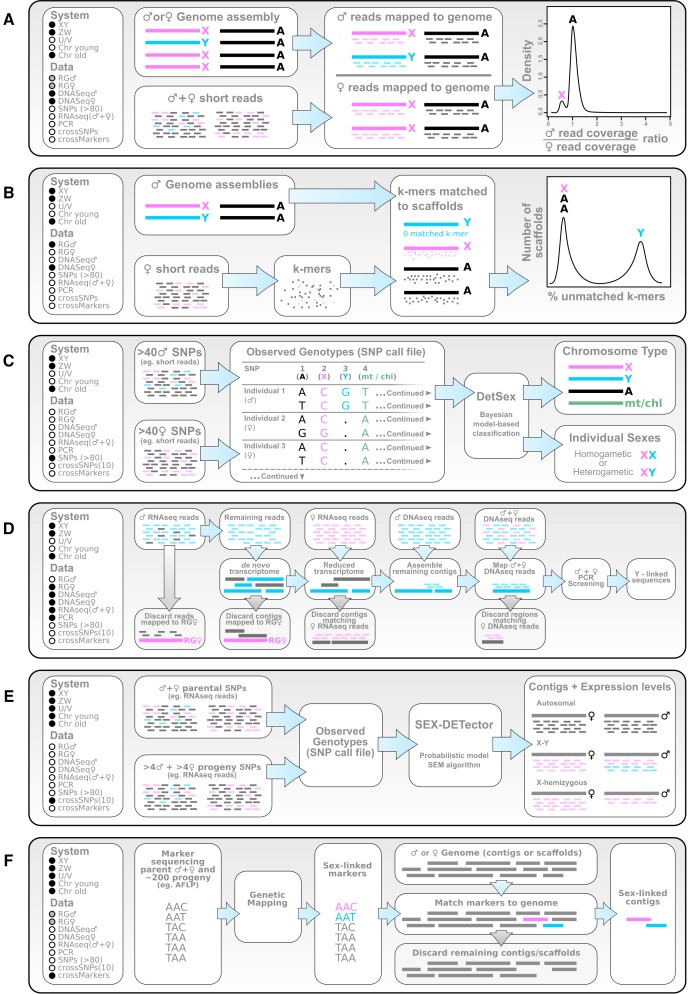
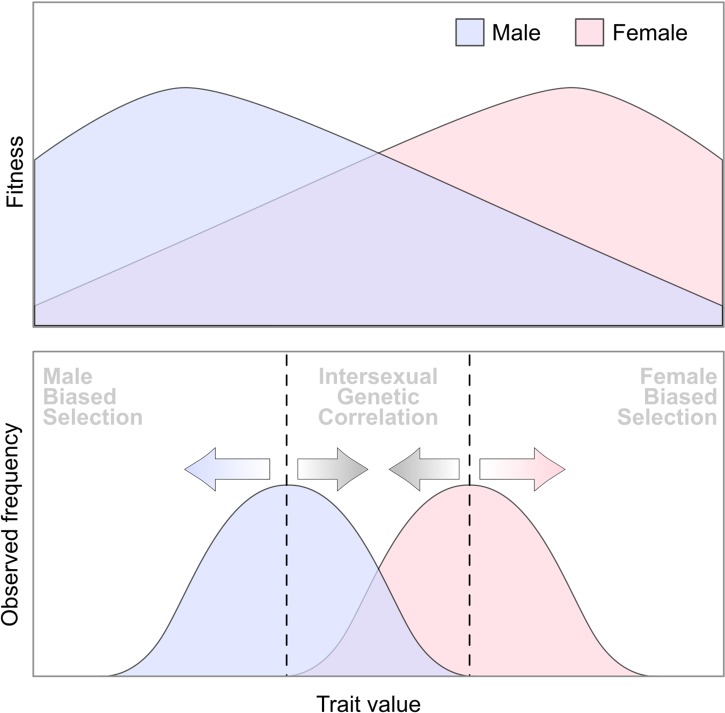
Plant sex chromosomes can be vastly different from those of the few historical animal model organisms from which most of our understanding of sex chromosome evolution is derived. Recently, we have seen several advancements from studies on green algae, brown algae, and land plants that are providing a broader understanding of the variable ways in which sex chromosomes can evolve in distant eukaryotic groups. Plant sex-determining genes are being identified and, as expected, are completely different from those in animals. Species with varying levels of differentiation between the X and Y have been found in plants, and these are hypothesized to be representing different stages of sex chromosome evolution. However, we are also finding that sex chromosomes can remain morphologically unchanged over extended periods of time. Where degeneration of the Y occurs, it appears to proceed similarly in plants and animals. Dosage compensation (a phenomenon that compensates for the consequent loss of expression from the Y) has now been documented in a plant system, its mechanism, however, remains unknown. Research has also begun on the role of sex chromosomes in sexual conflict resolution, and it appears that sex-biased genes evolve similarly in plants and animals, although the functions of these genes remain poorly studied. Because the difficulty in obtaining sex chromosome sequences is increasingly being overcome by methodological developments, there is great potential for further discovery within the field of plant sex chromosome evolution.
Keywords: Y degeneration; dioecy; sex chromosome sequencing; sex chromosome turnover; sex-biased expression.
© The Author(s) 2017. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Ahmed S, et al. 2014. A haploid system of sex determination in the brown alga Ectocarpus sp. Curr Biol. 24:1945–1957. - PubMed
-
- Akagi T, Henry IM, Tao R, Comai L. 2014. Plant genetics. A Y-chromosome-encoded small RNA acts as a sex determinant in persimmons. Science 346:646–650. - PubMed
-
- Al-Dous EK, et al. 2011. De novo genome sequencing and comparative genomics of date palm (Phoenix dactylifera). Nat Biotechnol. 29:521–527. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
