

Long-term, hormone-responsive organoid cultures of human endometrium in a chemically defined medium
- PMID: 28394884
- PMCID: PMC5410172
- DOI: 10.1038/ncb3516
Long-term, hormone-responsive organoid cultures of human endometrium in a chemically defined medium
Abstract
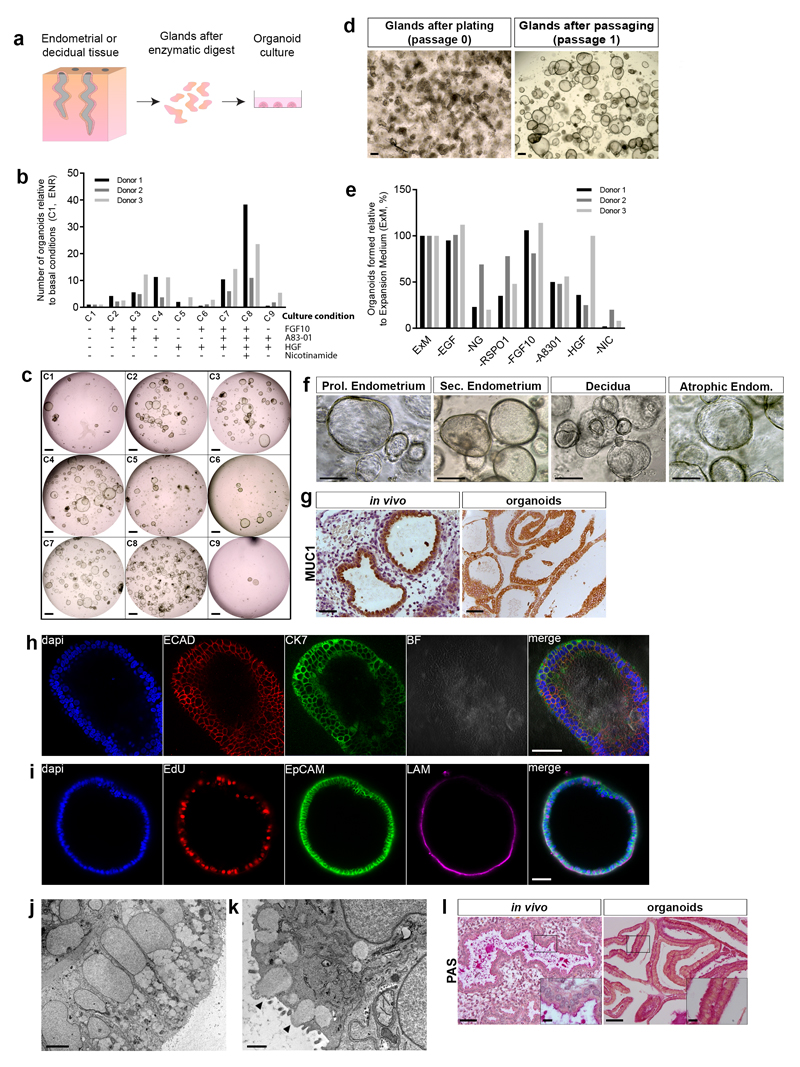
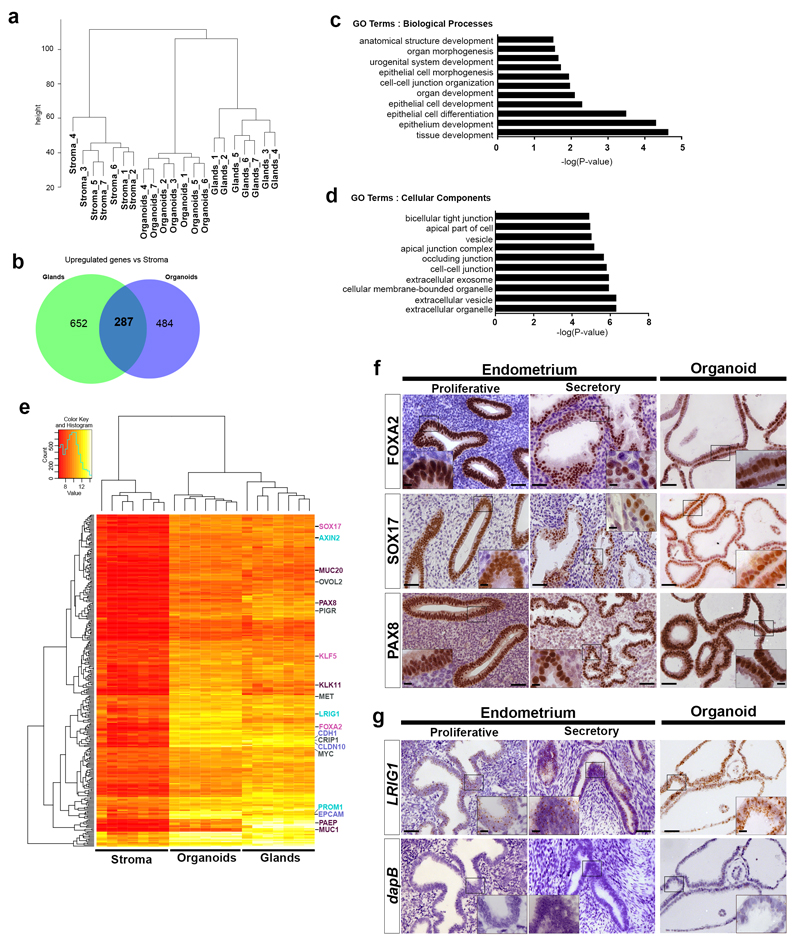
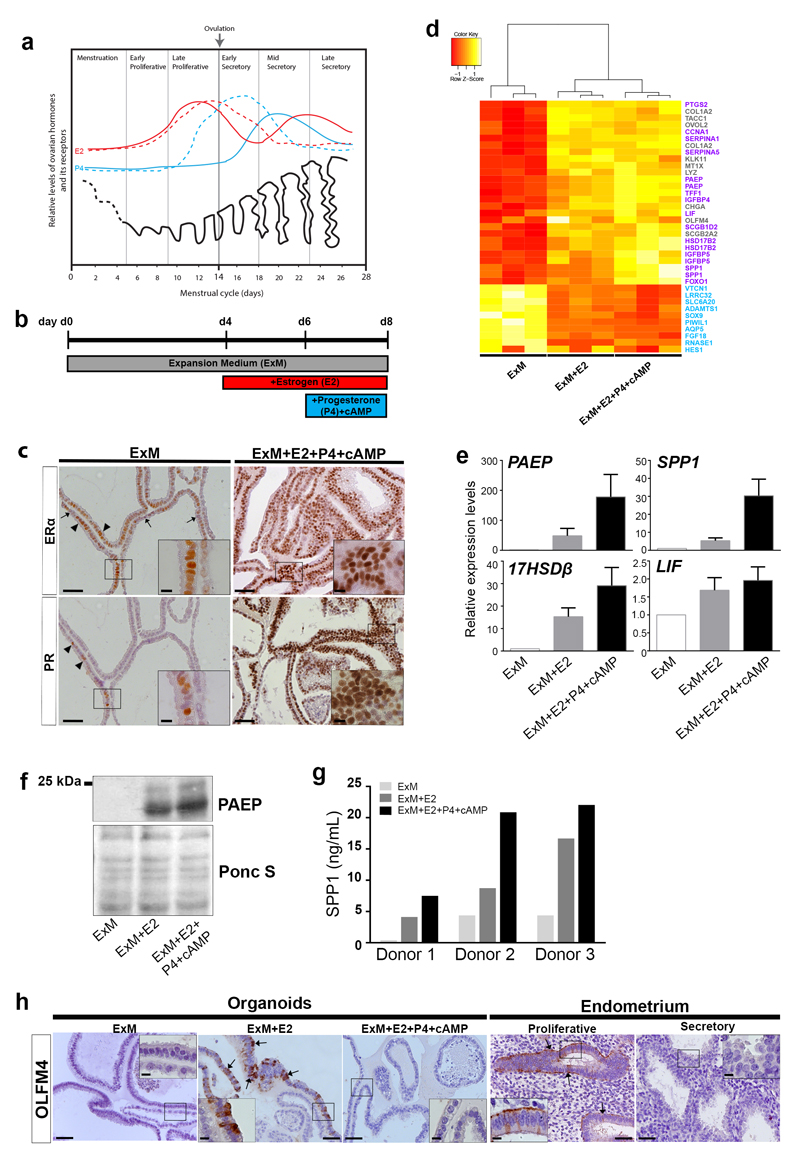
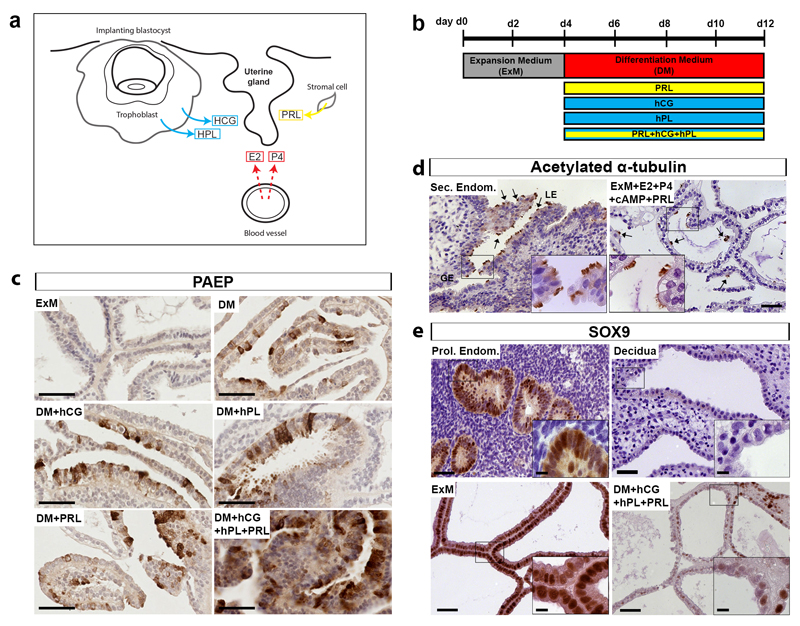
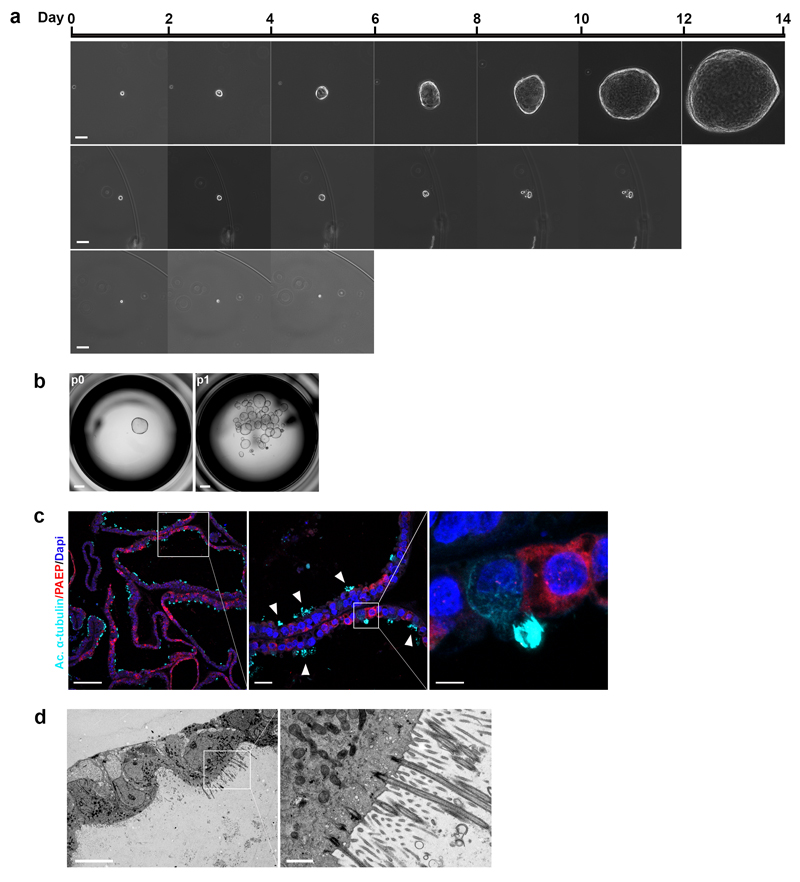
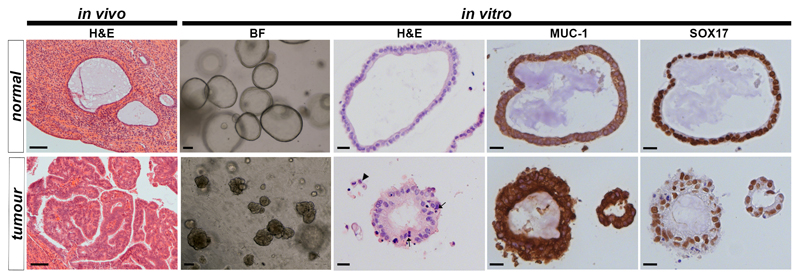
In humans, the endometrium, the uterine mucosal lining, undergoes dynamic changes throughout the menstrual cycle and pregnancy. Despite the importance of the endometrium as the site of implantation and nutritional support for the conceptus, there are no long-term culture systems that recapitulate endometrial function in vitro. We adapted conditions used to establish human adult stem-cell-derived organoid cultures to generate three-dimensional cultures of normal and decidualized human endometrium. These organoids expand long-term, are genetically stable and differentiate following treatment with reproductive hormones. Single cells from both endometrium and decidua can generate a fully functional organoid. Transcript analysis confirmed great similarity between organoids and the primary tissue of origin. On exposure to pregnancy signals, endometrial organoids develop characteristics of early pregnancy. We also derived organoids from malignant endometrium, and so provide a foundation to study common diseases, such as endometriosis and endometrial cancer, as well as the physiology of early gestation.
Figures
References
-
- Burton GJ, Watson AL, Hempstock J, Skepper JN, Jauniaux E. Uterine glands provide histiotrophic nutrition for the human fetus during the first trimester of pregnancy. The Journal of clinical endocrinology and metabolism. 2002;87:2954–2959. - PubMed
-
- Gray CA, Burghardt RC, Johnson GA, Bazer FW, Spencer TE. Evidence that absence of endometrial gland secretions in uterine gland knockout ewes compromises conceptus survival and elongation. Reproduction. 2002;124:289–300. - PubMed
-
- Filant J, Spencer TE. Endometrial glands are essential for blastocyst implantation and decidualization in the mouse uterus. Biology of reproduction. 2013;88:93. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
