

Exploring binaural hearing in gerbils (Meriones unguiculatus) using virtual headphones
- PMID: 28394906
- PMCID: PMC5386270
- DOI: 10.1371/journal.pone.0175142
Exploring binaural hearing in gerbils (Meriones unguiculatus) using virtual headphones
Abstract
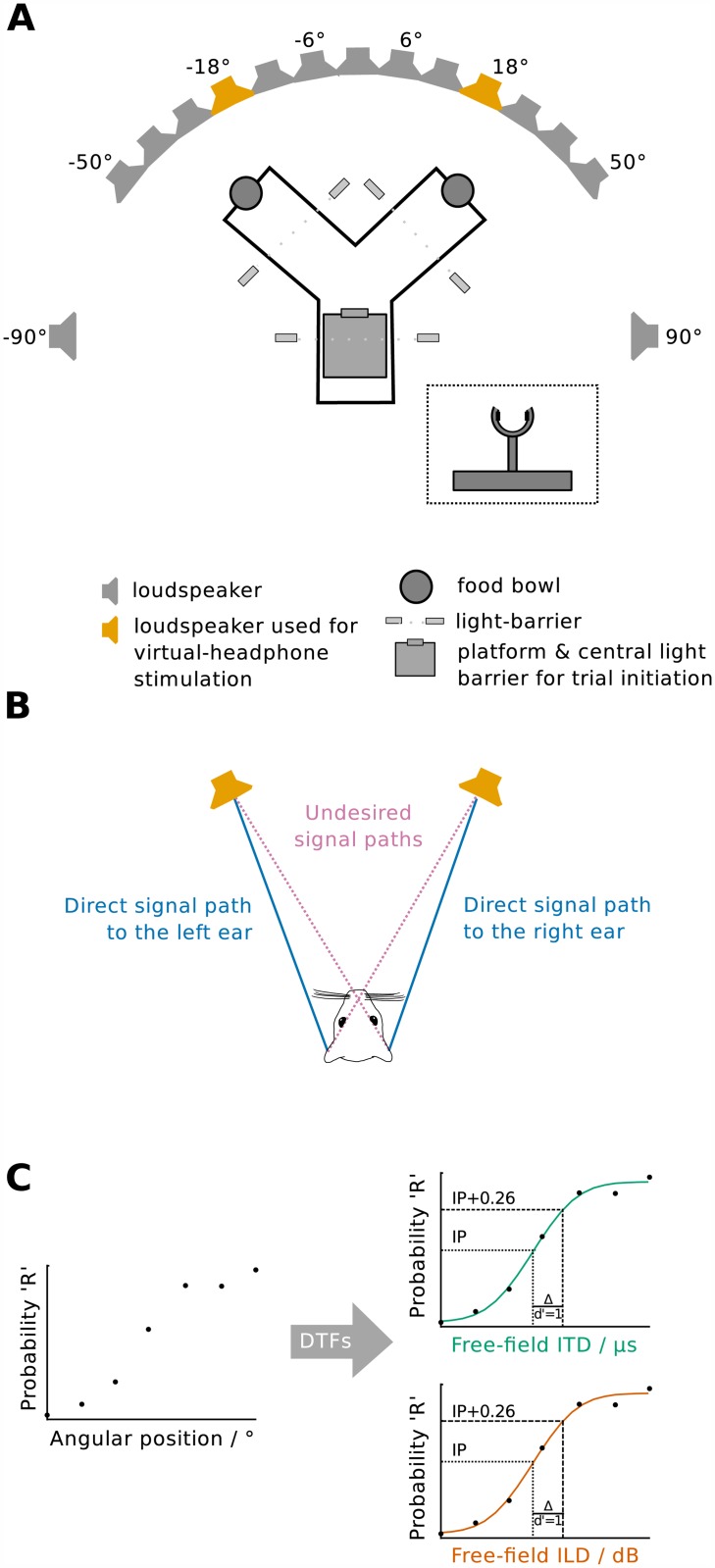
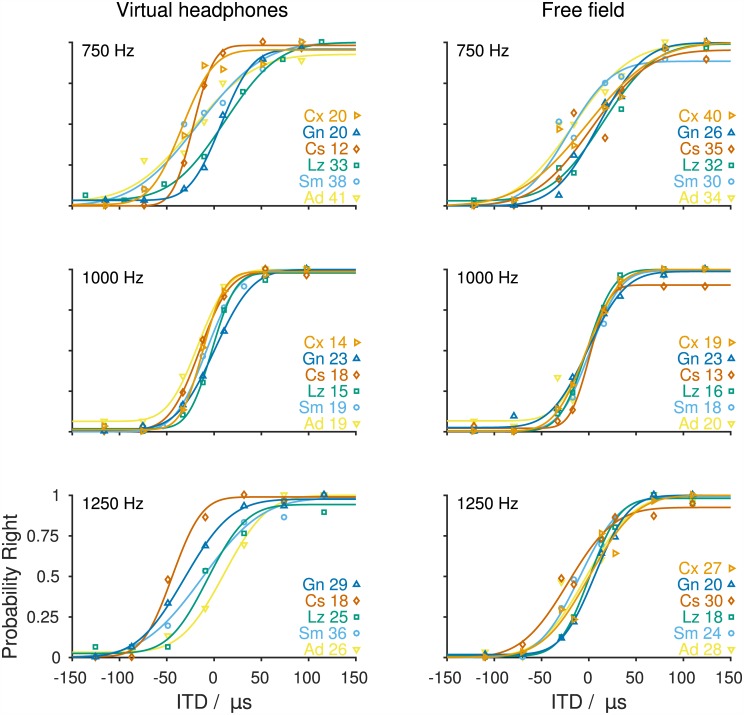
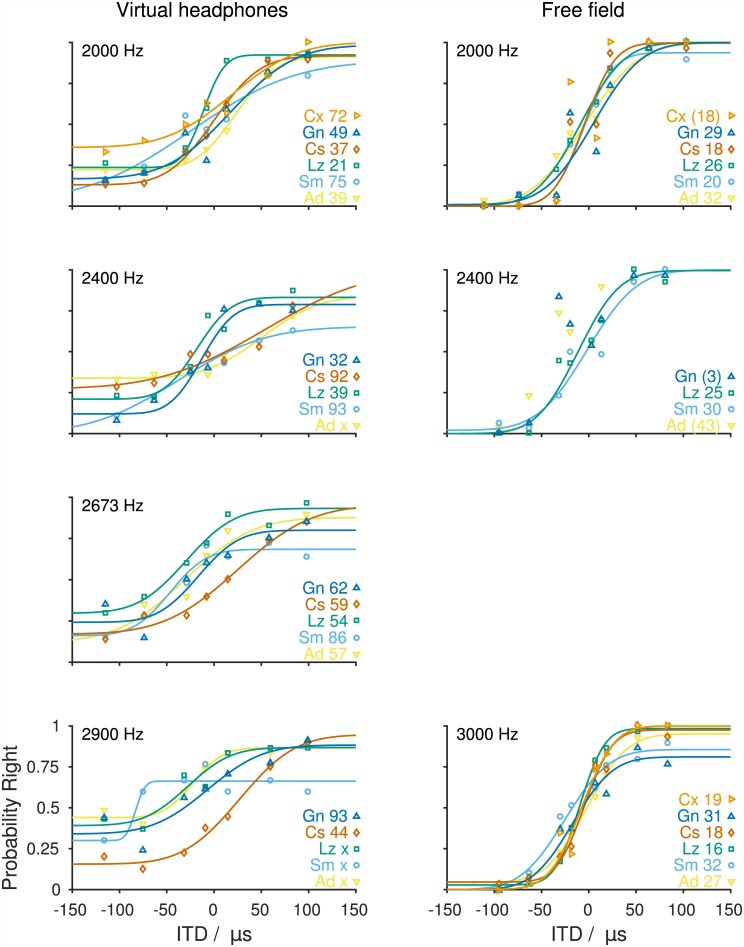
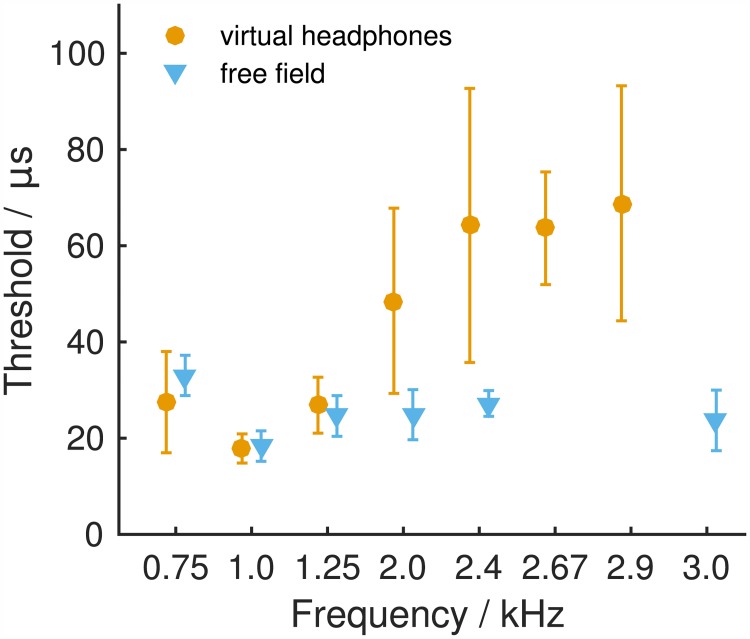
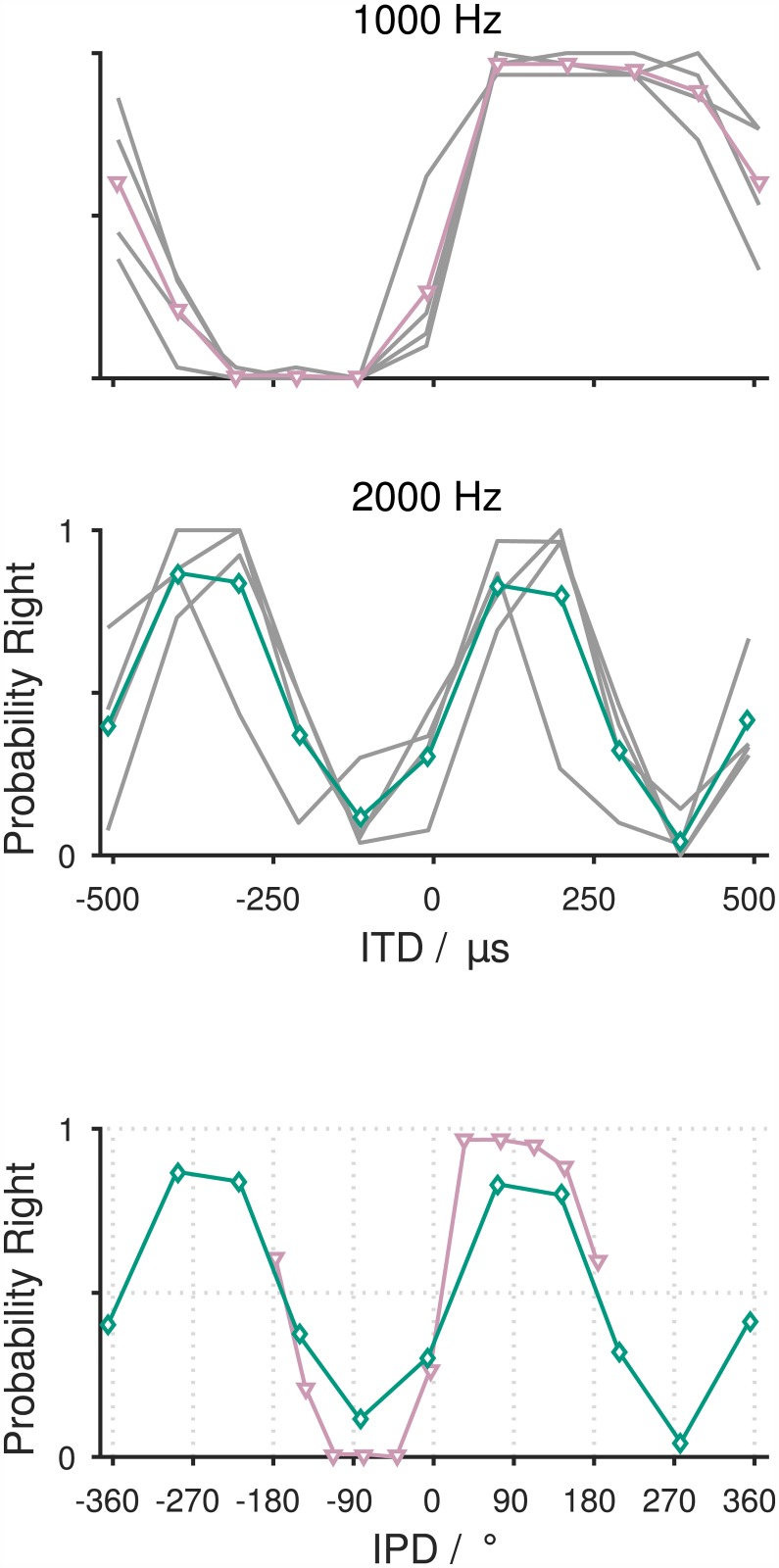
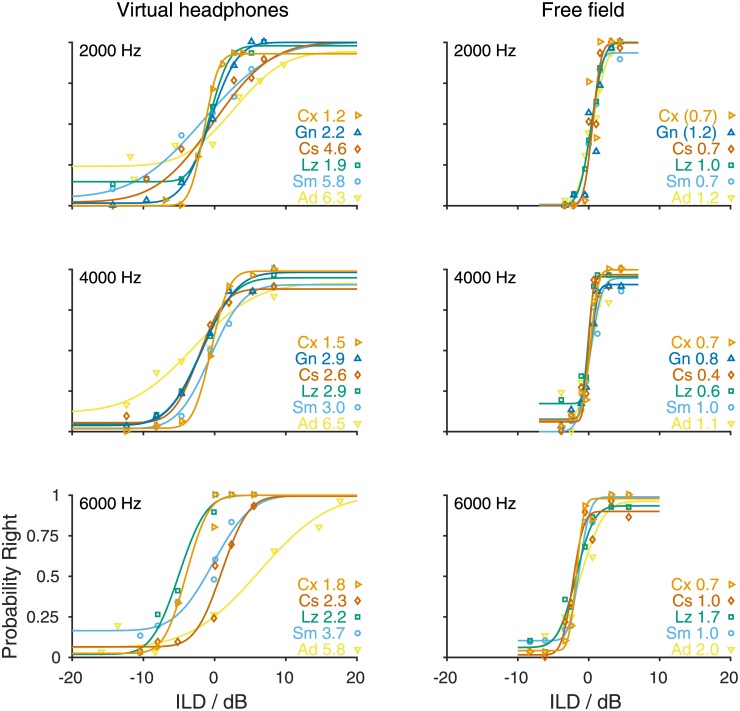
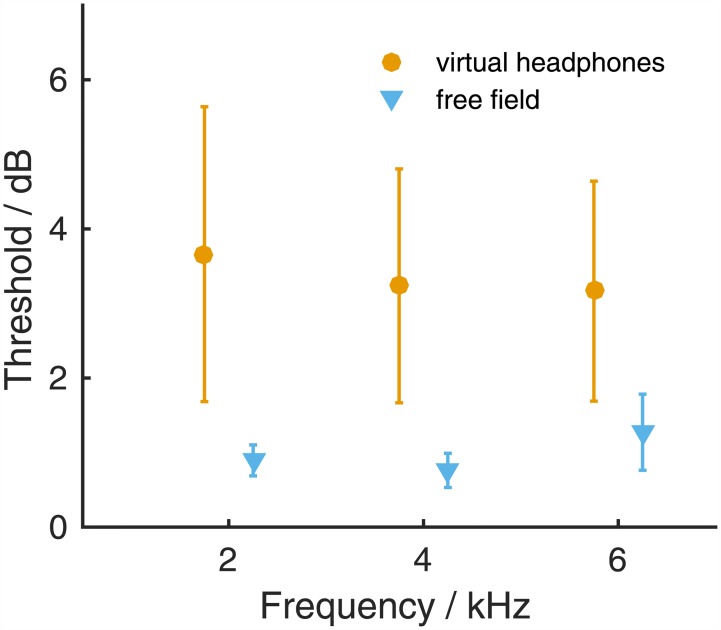
The Mongolian gerbil (Meriones unguiculatus) has become a key species in investigations of the neural processing of sound localization cues in mammals. While its sound localization has been tested extensively under free-field stimulation, many neurophysiological studies use headphones to present signals with binaural localization cues. The gerbil's behavioral sensitivity to binaural cues, however, is unknown for the lack of appropriate stimulation paradigms in awake behaving gerbils. We close this gap in knowledge by mimicking a headphone stimulation; we use free-field loudspeakers and apply cross-talk cancellation techniques to present pure tones with binaural cues via "virtual headphones" to gerbils trained in a sound localization task. All gerbils were able to lateralize sounds depending on the interaural time or level difference (ITD and ILD, respectively). For ITD stimuli, reliable responses were seen for frequencies ≤2.9 kHz, the highest frequency tested with ITD stimuli. ITD sensitivity was frequency-dependent with the highest sensitivity observed at 1 kHz. For stimuli with ITD outside the gerbil's physiological range, responses were cyclic indicating the use of phase information when lateralizing narrow-band sounds. For ILD stimuli, reliable responses were obtained for frequencies ≥2 kHz. The comparison of ITD and ILD thresholds with ITD and ILD thresholds derived from gerbils' free-field performance suggests that ongoing ITD information is the main cue for sound localization at frequencies <2 kHz. At 2 kHz, ITD and ILD cues are likely used in a complementary way. Verification of the use of the virtual headphones suggests that they can serve as a suitable substitute for conventional headphones particularly at frequencies ≤2 kHz.
Conflict of interest statement
Figures
References
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
