

Selective entrainment of gamma subbands by different slow network oscillations
- PMID: 28396398
- PMCID: PMC5410835
- DOI: 10.1073/pnas.1617249114
Selective entrainment of gamma subbands by different slow network oscillations
Abstract
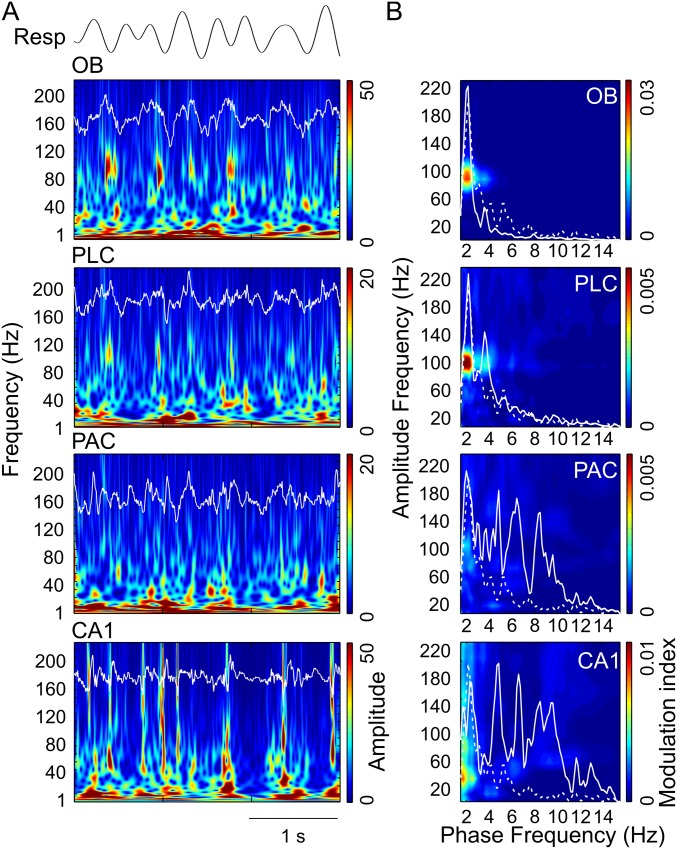
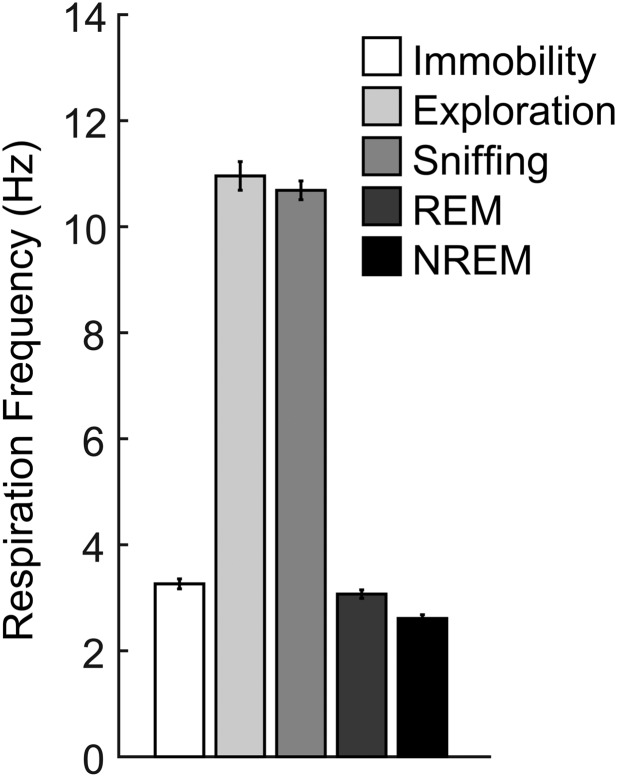
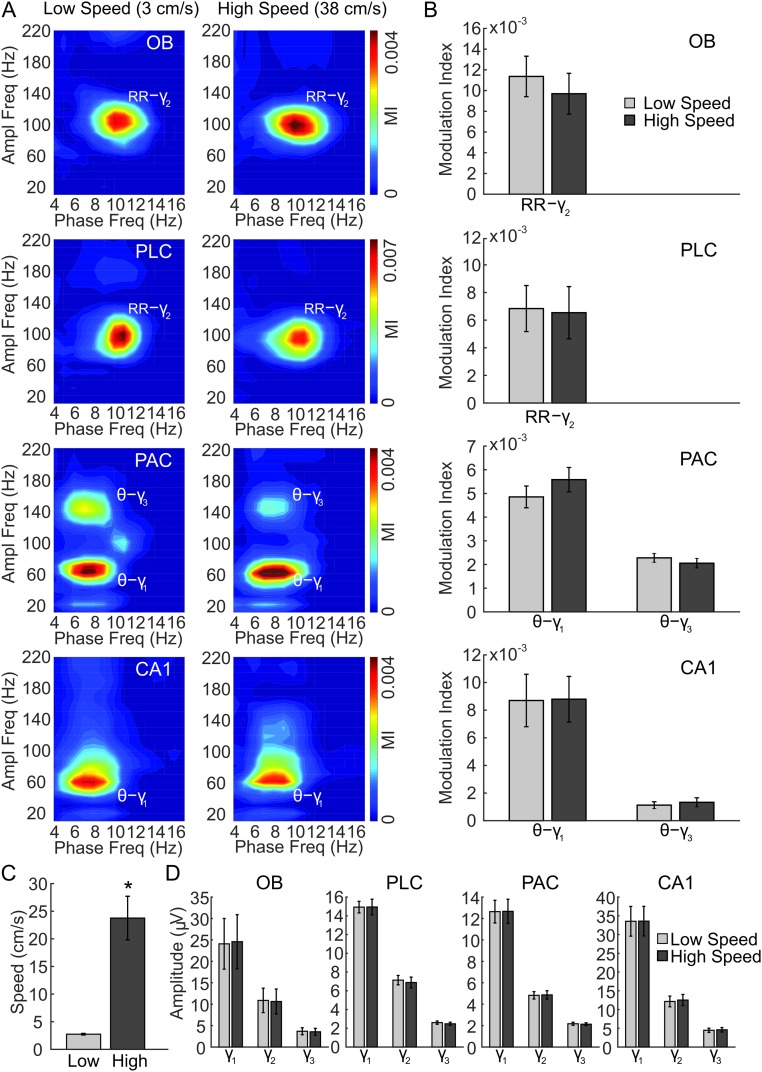
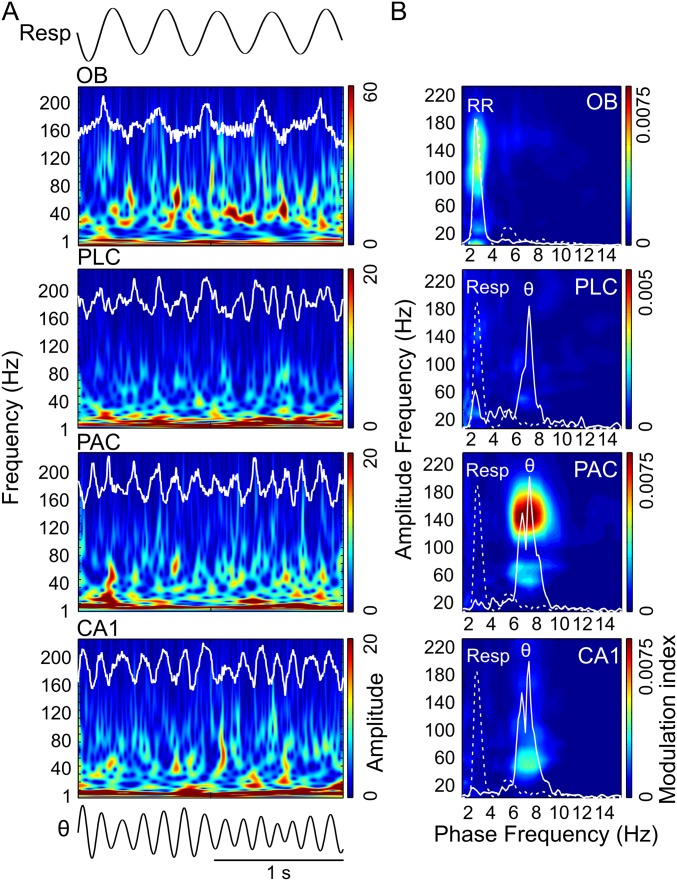
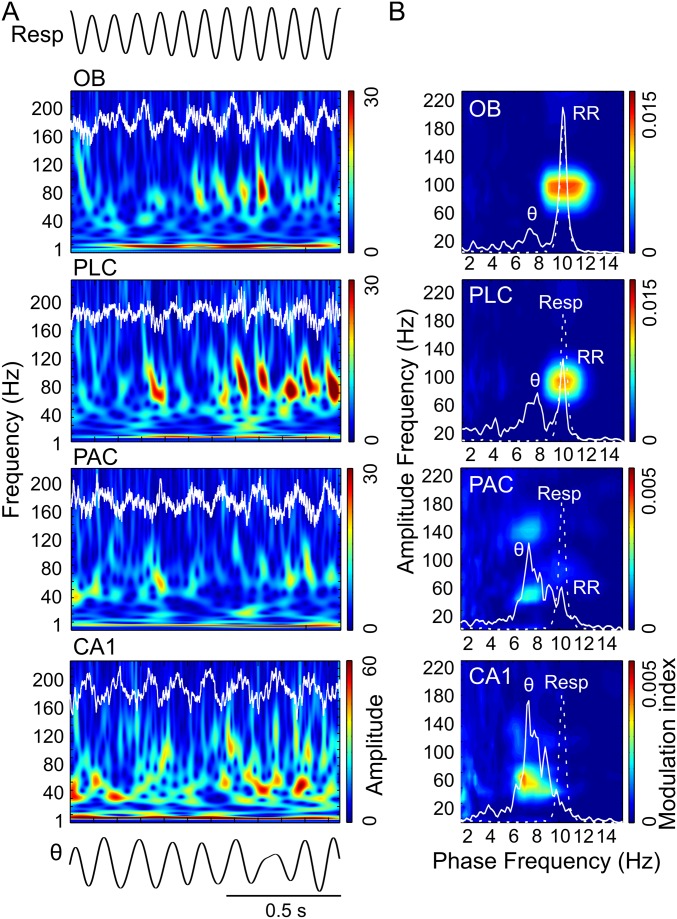
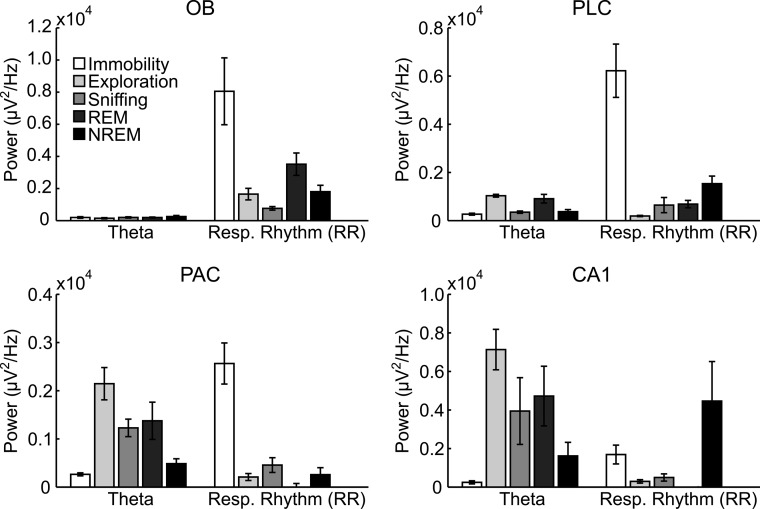
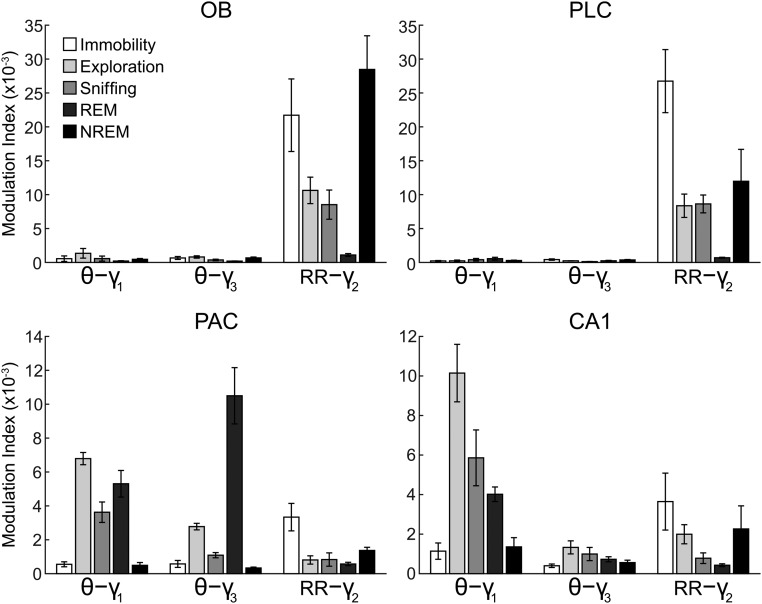
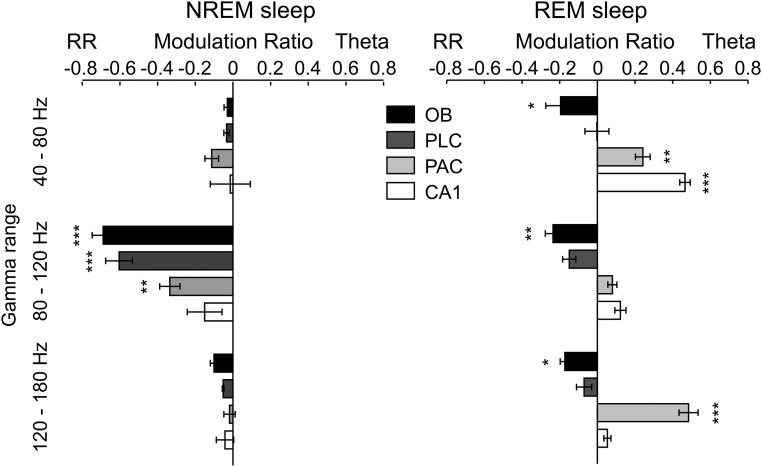
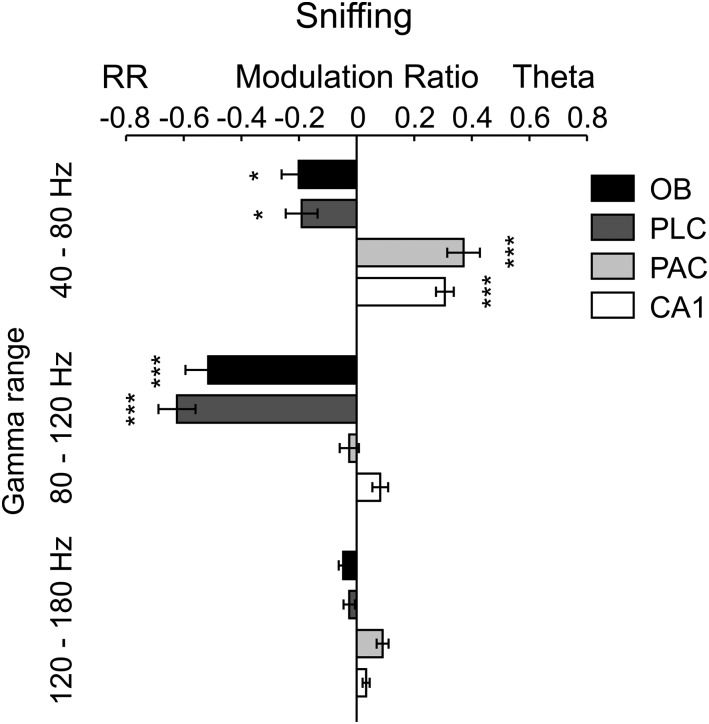
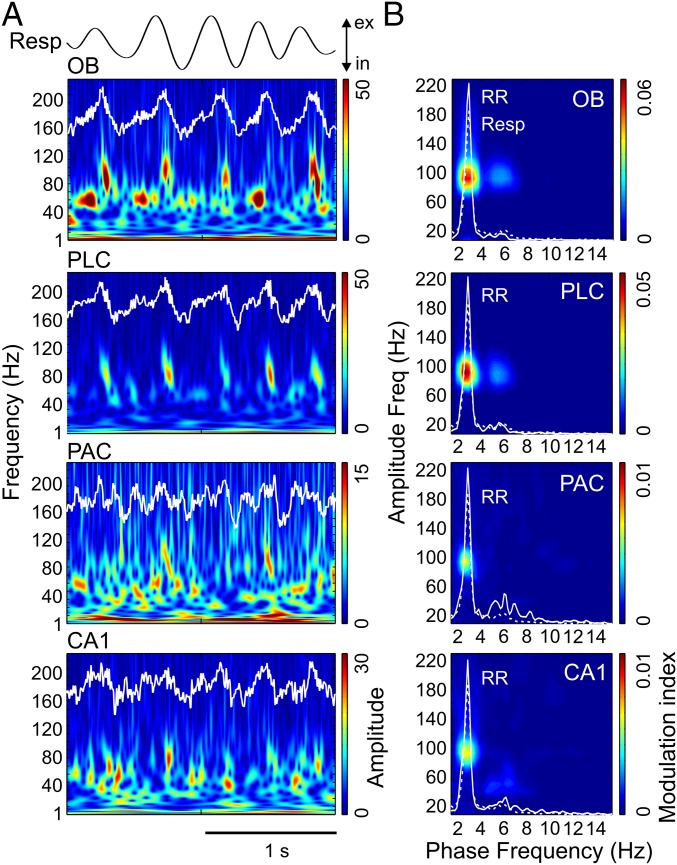
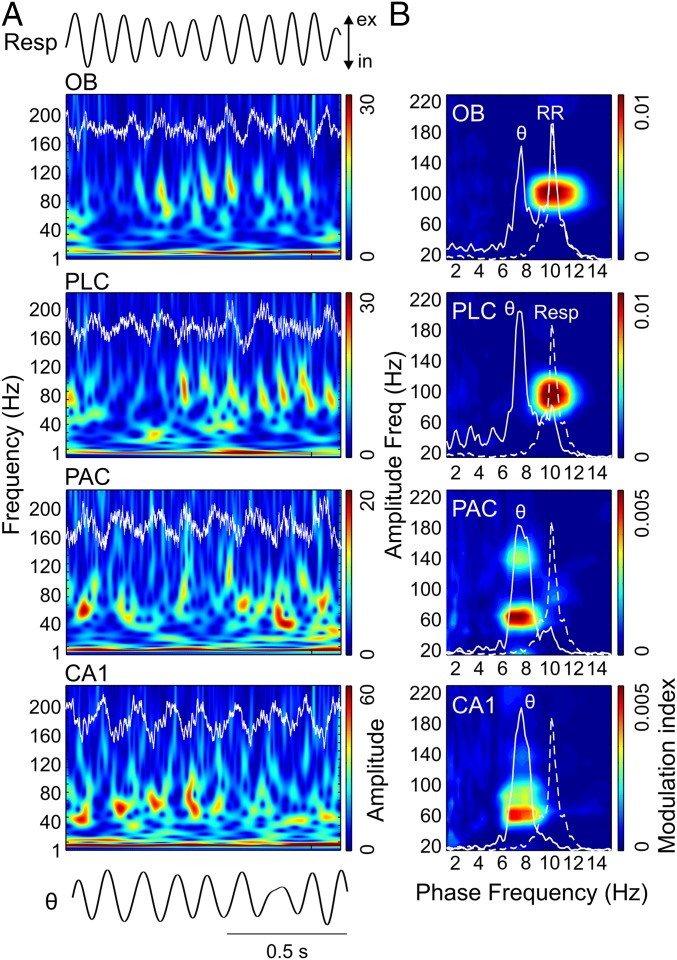
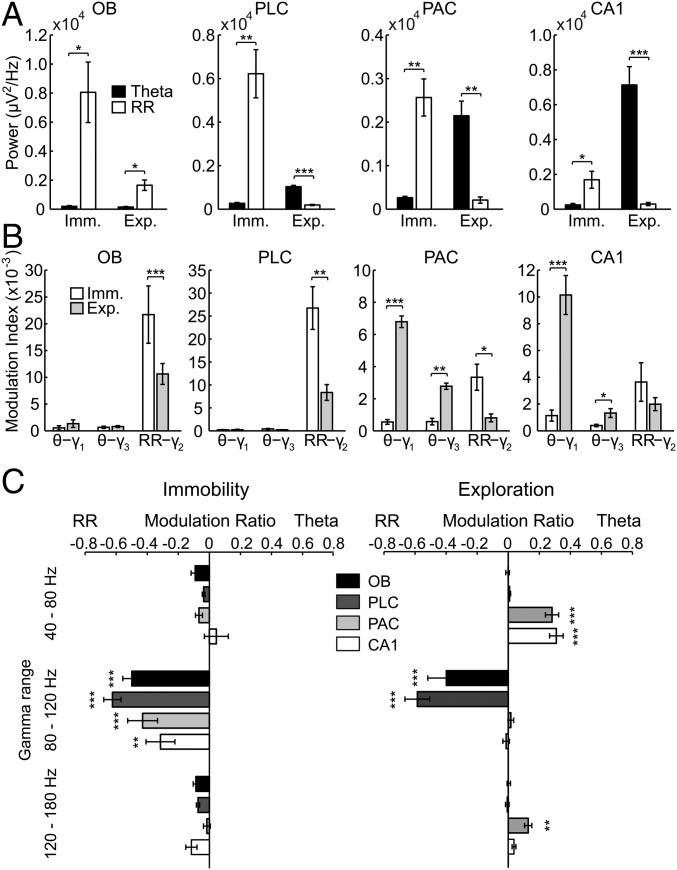
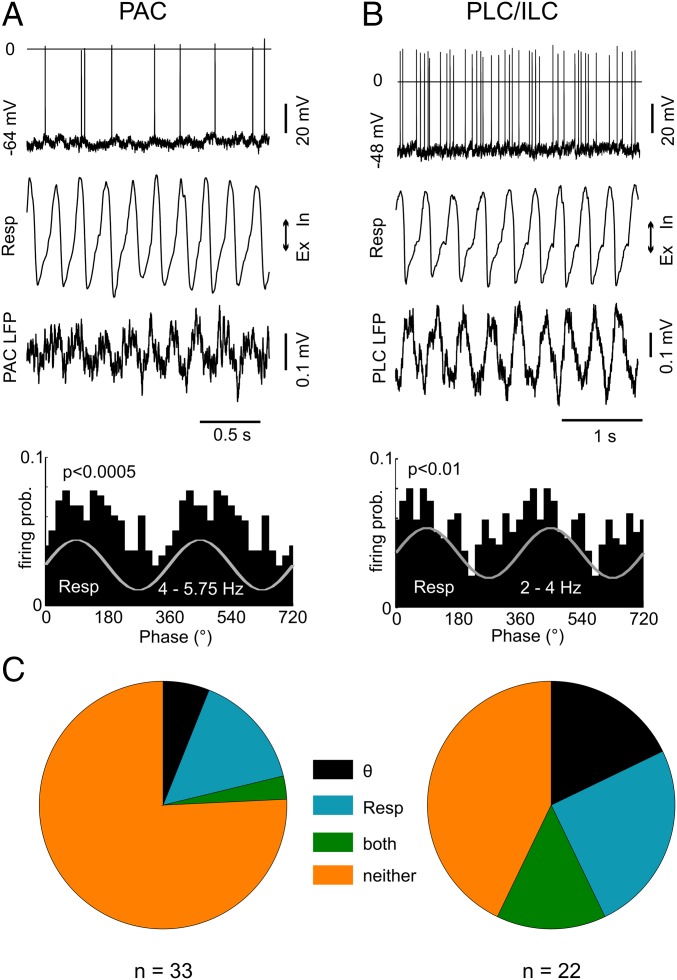
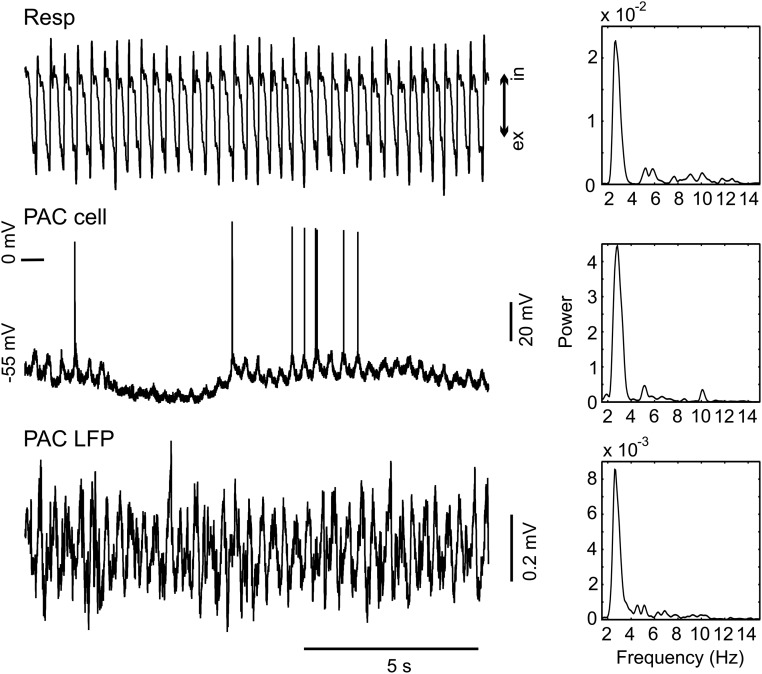
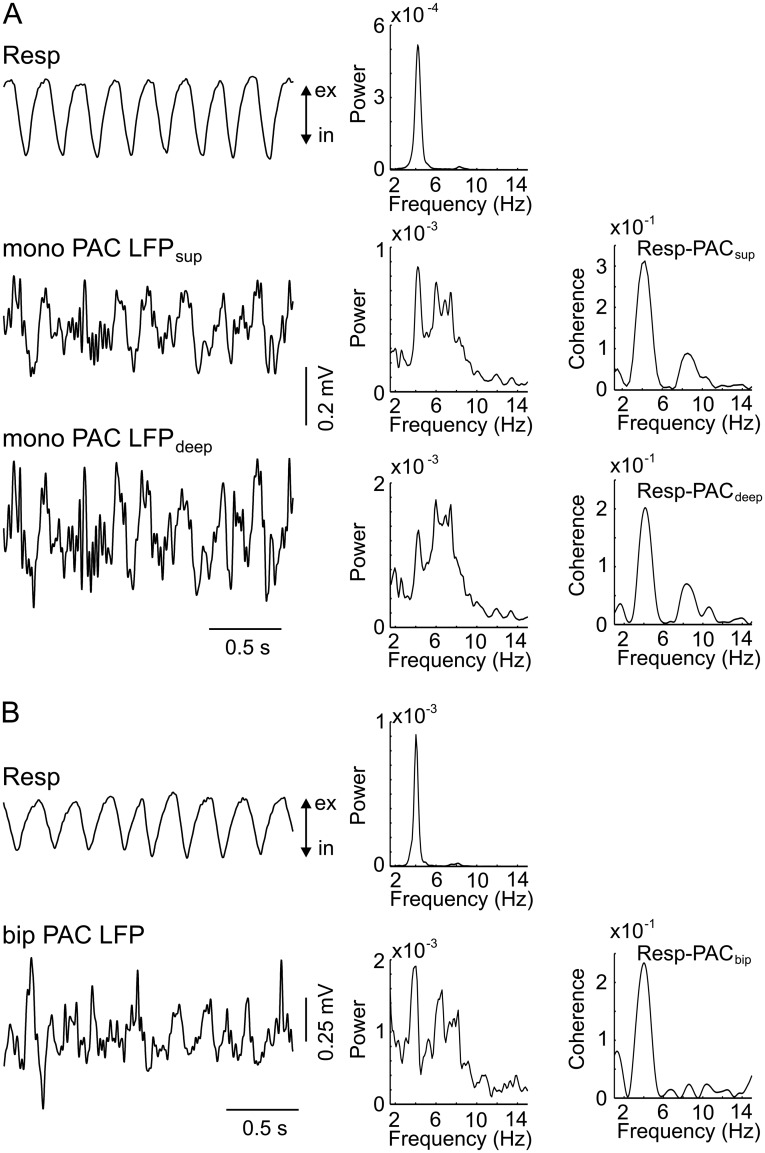
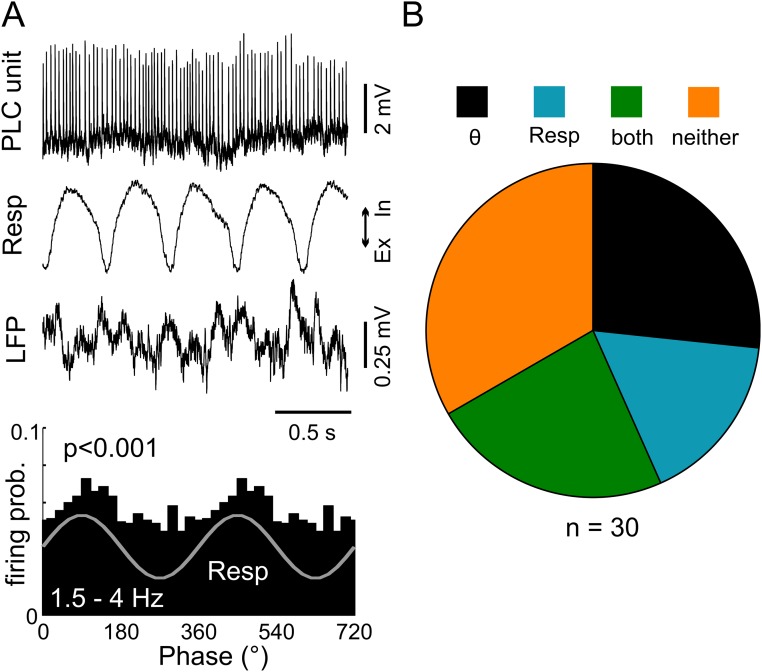
Theta oscillations (4-12 Hz) are thought to provide a common temporal reference for the exchange of information among distant brain networks. On the other hand, faster gamma-frequency oscillations (30-160 Hz) nested within theta cycles are believed to underlie local information processing. Whether oscillatory coupling between global and local oscillations, as showcased by theta-gamma coupling, is a general coding mechanism remains unknown. Here, we investigated two different patterns of oscillatory network activity, theta and respiration-induced network rhythms, in four brain regions of freely moving mice: olfactory bulb (OB), prelimbic cortex (PLC), parietal cortex (PAC), and dorsal hippocampus [cornu ammonis 1 (CA1)]. We report differential state- and region-specific coupling between the slow large-scale rhythms and superimposed fast oscillations. During awake immobility, all four regions displayed a respiration-entrained rhythm (RR) with decreasing power from OB to CA1, which coupled exclusively to the 80- to 120-Hz gamma subband (γ2). During exploration, when theta activity was prevailing, OB and PLC still showed exclusive coupling of RR with γ2 and no theta-gamma coupling, whereas PAC and CA1 switched to selective coupling of theta with 40- to 80-Hz (γ1) and 120- to 160-Hz (γ3) gamma subbands. Our data illustrate a strong, specific interaction between neuronal activity patterns and respiration. Moreover, our results suggest that the coupling between slow and fast oscillations is a general brain mechanism not limited to the theta rhythm.
Keywords: cross-frequency coupling; gamma subbands; neocortex; respiration; theta.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
