

Structural hierarchy controlling dimerization and target DNA recognition in the AHR transcriptional complex
- PMID: 28396409
- PMCID: PMC5448172
- DOI: 10.1073/pnas.1617035114
Structural hierarchy controlling dimerization and target DNA recognition in the AHR transcriptional complex
Abstract
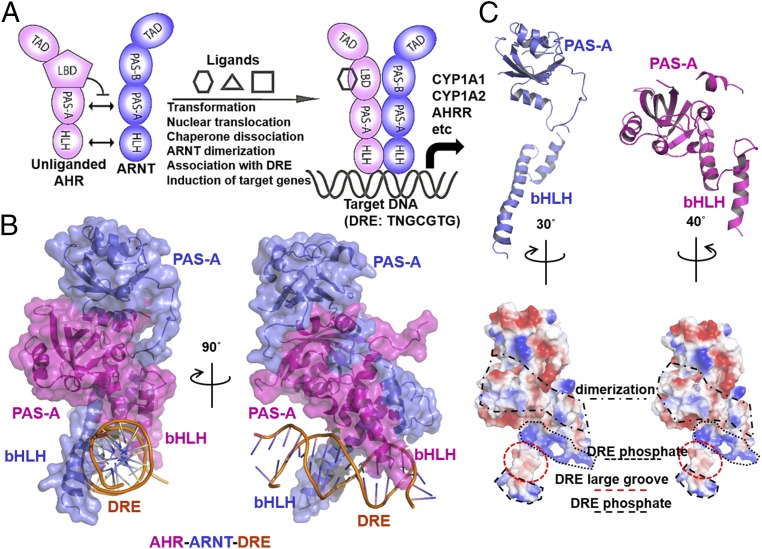
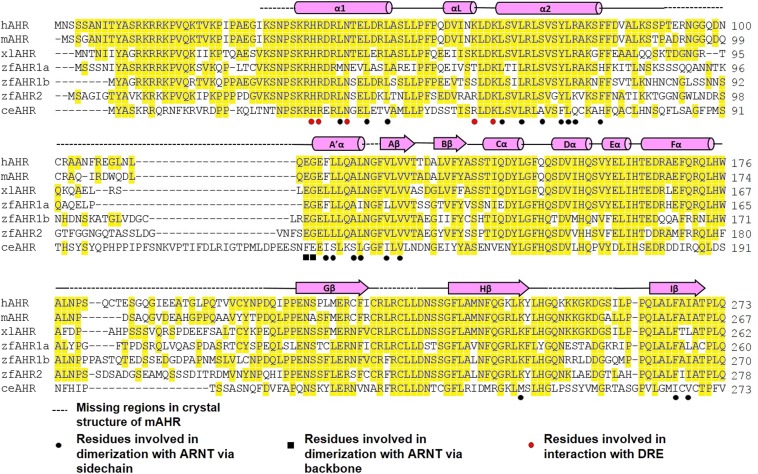
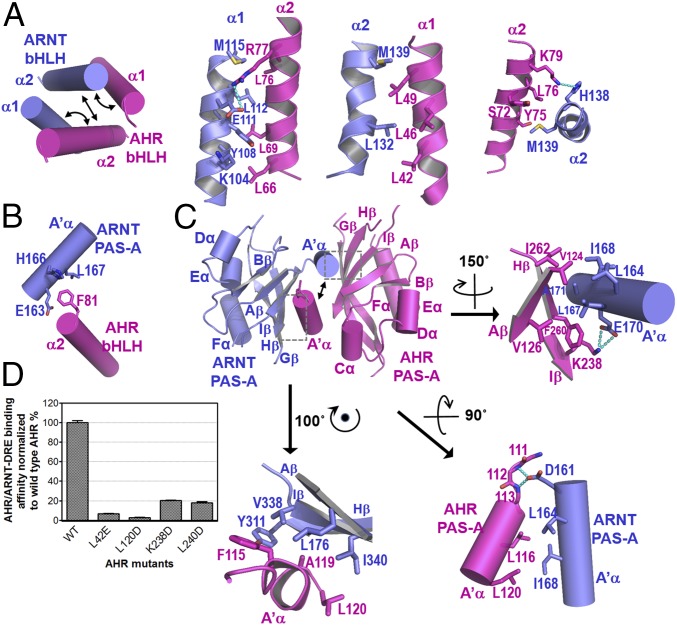
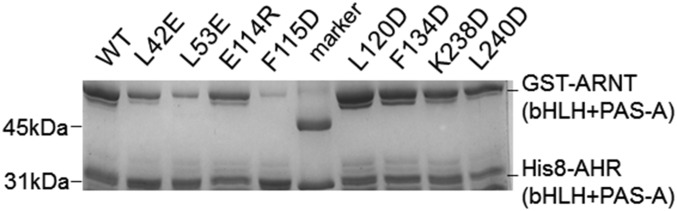
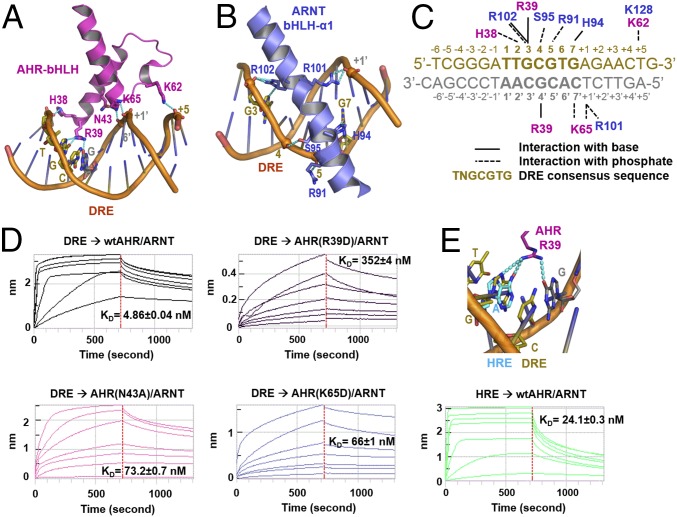
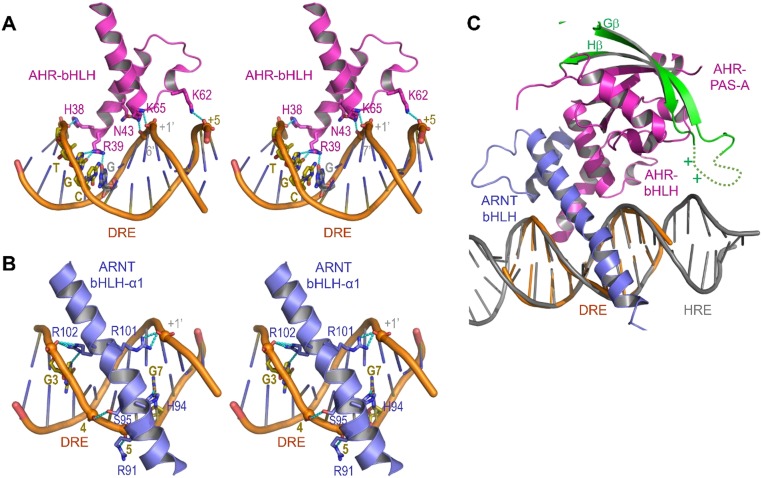
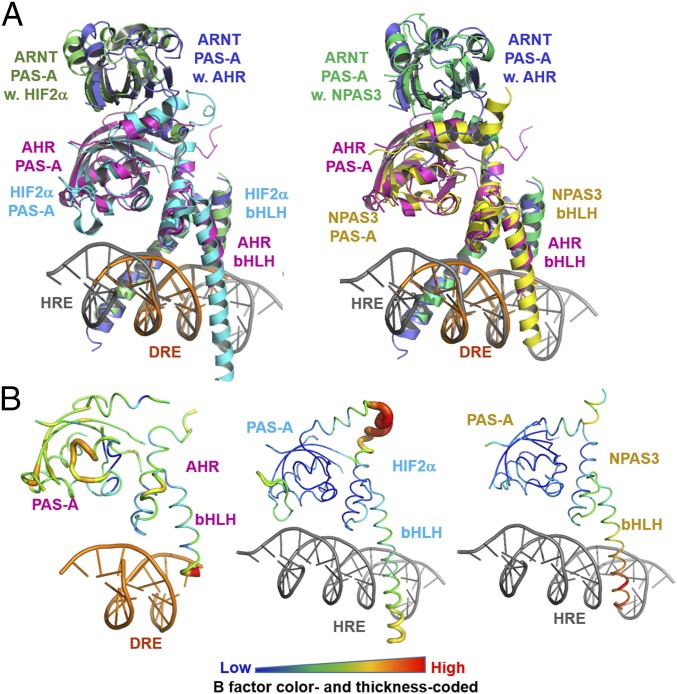
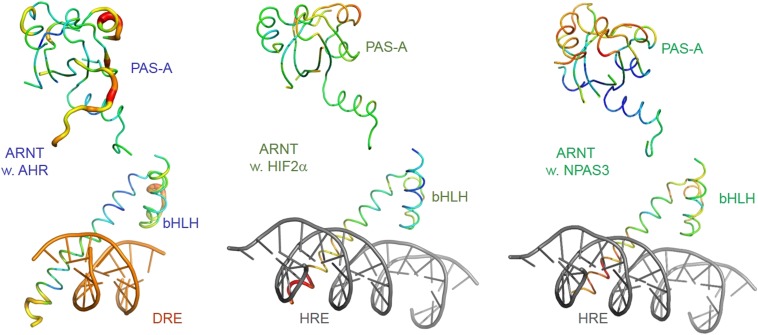
The aryl hydrocarbon receptor (AHR) belongs to the PAS (PER-ARNT-SIM) family transcription factors and mediates broad responses to numerous environmental pollutants and cellular metabolites, modulating diverse biological processes from adaptive metabolism, acute toxicity, to normal physiology of vascular and immune systems. The AHR forms a transcriptionally active heterodimer with ARNT (AHR nuclear translocator), which recognizes the dioxin response element (DRE) in the promoter of downstream genes. We determined the crystal structure of the mammalian AHR-ARNT heterodimer in complex with the DRE, in which ARNT curls around AHR into a highly intertwined asymmetric architecture, with extensive heterodimerization interfaces and AHR interdomain interactions. Specific recognition of the DRE is determined locally by the DNA-binding residues, which discriminates it from the closely related hypoxia response element (HRE), and is globally affected by the dimerization interfaces and interdomain interactions. Changes at the interdomain interactions caused either AHR constitutive nuclear localization or failure to translocate to nucleus, underlying an allosteric structural pathway for mediating ligand-induced exposure of nuclear localization signal. These observations, together with the global higher flexibility of the AHR PAS-A and its loosely packed structural elements, suggest a dynamic structural hierarchy for complex scenarios of AHR activation induced by its diverse ligands.
Keywords: AHR; ARNT; DNA recognition; dimerization; transcriptional complex.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Assembly and function of bHLH-PAS complexes.Proc Natl Acad Sci U S A. 2017 May 23;114(21):5330-5332. doi: 10.1073/pnas.1705408114. Epub 2017 May 15. Proc Natl Acad Sci U S A. 2017. PMID: 28507151 Free PMC article. No abstract available.
References
-
- Schmidt JV, Bradfield CA. Ah receptor signaling pathways. Annu Rev Cell Dev Biol. 1996;12:55–89. - PubMed
-
- Thomas PE, Hutton JJ. Genetics of aryl hydrocarbon hydroxylase induction in mice: Additive inheritance in crosses between C3H-HeJ and DBA-2J. Biochem Genet. 1973;8:249–257. - PubMed
-
- Thomas PE, Kouri RE, Hutton JJ. The genetics of aryl hydrocarbon hydroxylase induction in mice: A single gene difference between C57BL-6J and DBA-2J. Biochem Genet. 1972;6:157–168. - PubMed
-
- Poland AP, Glover E, Robinson JR, Nebert DW. Genetic expression of aryl hydrocarbon hydroxylase activity. Induction of monooxygenase activities and cytochrome P1-450 formation by 2,3,7,8-tetrachlorodibenzo-p-dioxin in mice genetically “nonresponsive” to other aromatic hydrocarbons. J Biol Chem. 1974;249:5599–5606. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
