

Pregnancy Reprograms Large-Conductance Ca2+-Activated K+ Channel in Uterine Arteries: Roles of Ten-Eleven Translocation Methylcytosine Dioxygenase 1-Mediated Active Demethylation
- PMID: 28396535
- PMCID: PMC5426984
- DOI: 10.1161/HYPERTENSIONAHA.117.09059
Pregnancy Reprograms Large-Conductance Ca2+-Activated K+ Channel in Uterine Arteries: Roles of Ten-Eleven Translocation Methylcytosine Dioxygenase 1-Mediated Active Demethylation
Abstract
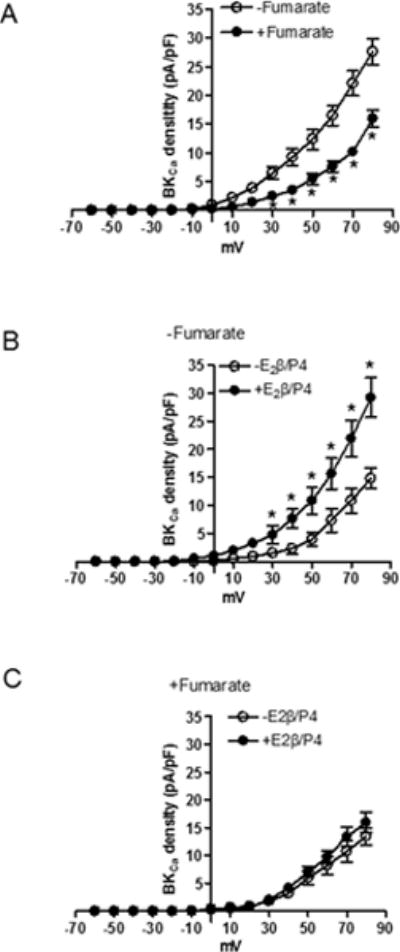
The large-conductance Ca2+-activated K+ (BKCa) channel is of critical importance in pregnancy-mediated increase in uterine artery vasodilation and blood flow. The present study tested the hypothesis that active DNA demethylation plays a key role in pregnancy-induced reprogramming and upregulation of BKCa channel β1 subunit (BKβ1) in uterine arteries. Uterine arteries were isolated from nonpregnant and near-term pregnant sheep. Pregnancy significantly increased the expression of ten-eleven translocation methylcytosine dioxygenase 1 (TET1) in uterine arteries. A half-palindromic estrogen response element was identified at the TET1 promoter, and estrogen treatment increased TET1 promoter activity and TET1 expression in uterine arteries. In accordance, pregnancy and steroid hormone treatment resulted in demethylation of BKβ1 promoter by increasing 5-hydroxymethylcytosine and decreasing 5-methylcytosine at the CpG in the Sp1-380 binding site that is of critical importance in the regulation of the promoter activity and BKβ1 expression. Inhibition of TET1 with fumarate significantly decreased BKβ1 expression in uterine arteries of pregnant animals and blocked steroid hormone-induced upregulation of BKβ1. Functionally, fumarate treatment inhibited pregnancy and steroid hormone-induced increases in BKCa channel current density and BKCa channel-mediated relaxations. In addition, fumarate blocked pregnancy and steroid hormone-induced decrease in pressure-dependent myogenic tone of the uterine artery. The results demonstrate a novel mechanism of estrogen-mediated active DNA demethylation in reprogramming of BKCa channel expression and function in the adaption of uterine circulation during pregnancy.
Keywords: 5-hydroxymethylcytosine; 5-methylcytosine; RNA, small interfering; estrogens; pregnancy.
© 2017 American Heart Association, Inc.
Figures





References
-
- Ledoux J, Werner ME, Brayden JE, Nelson MT. Calcium-activated potassium channels and the regulation of vascular tone. Physiology. 2006;21:69–78. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
