

Salicylidene Acylhydrazides and Hydroxyquinolines Act as Inhibitors of Type Three Secretion Systems in Pseudomonas aeruginosa by Distinct Mechanisms
- PMID: 28396545
- PMCID: PMC5444141
- DOI: 10.1128/AAC.02566-16
Salicylidene Acylhydrazides and Hydroxyquinolines Act as Inhibitors of Type Three Secretion Systems in Pseudomonas aeruginosa by Distinct Mechanisms
Abstract
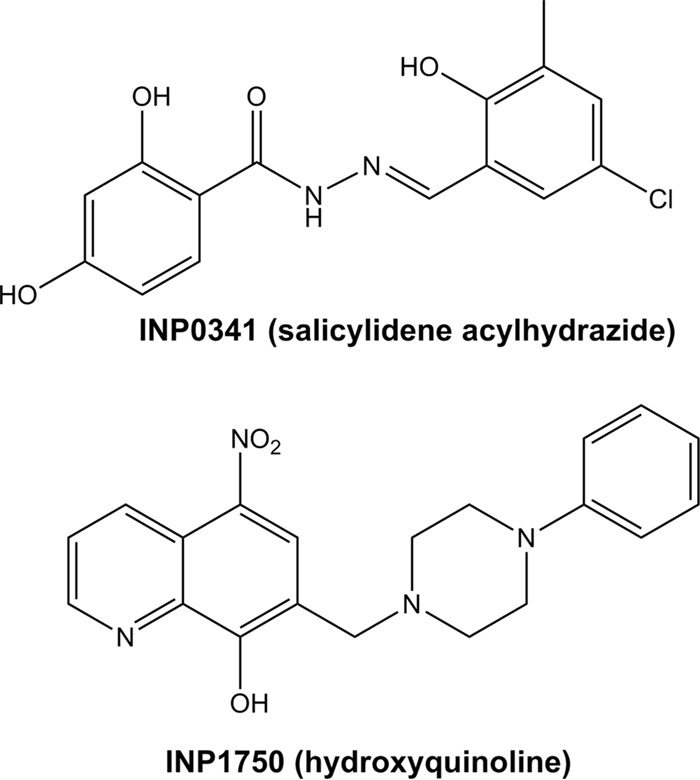
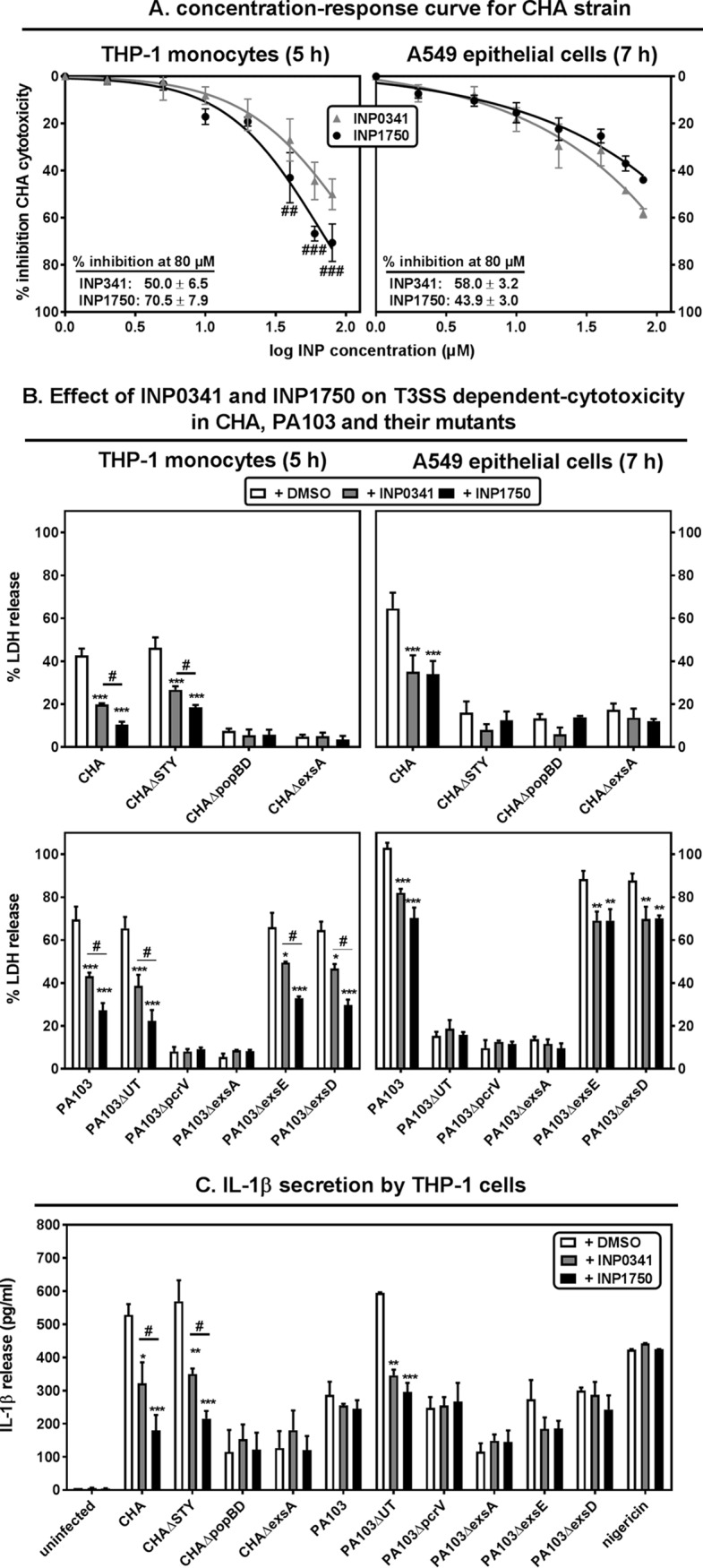
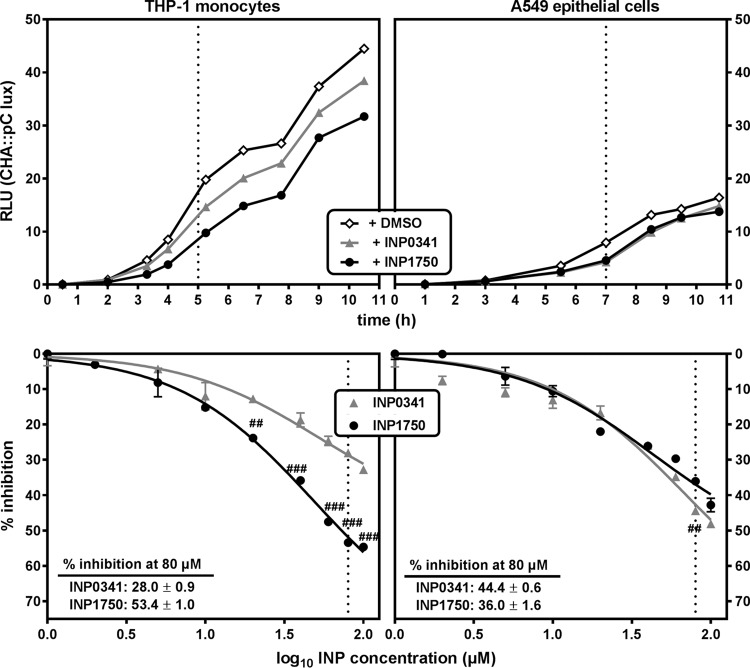
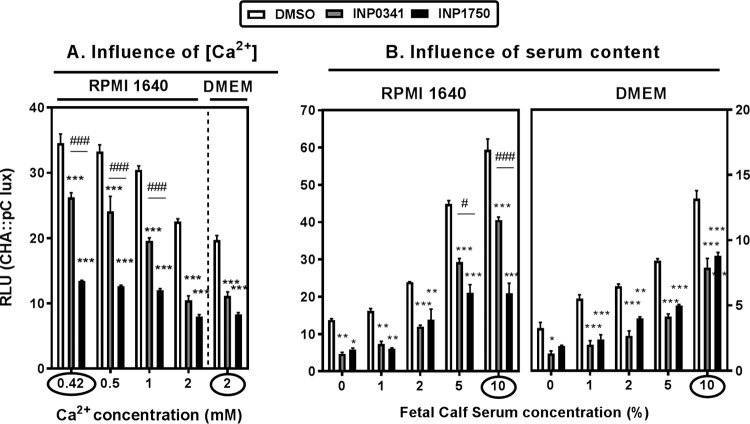
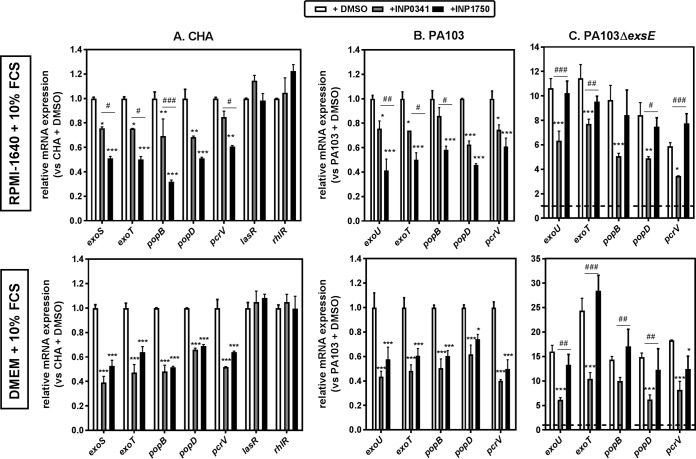
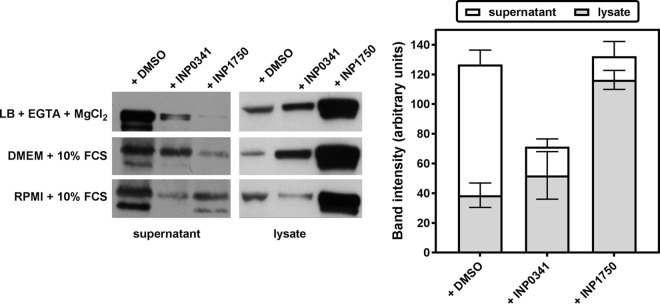
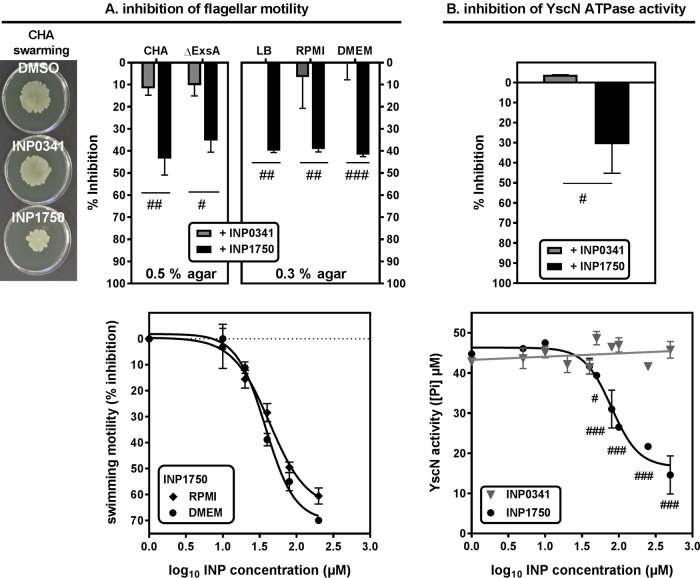
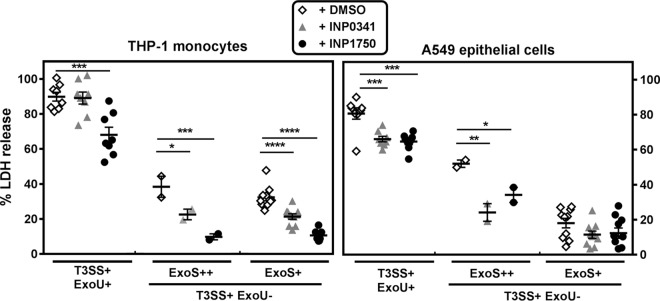
Type 3 secretion systems (T3SSs) are major virulence factors in Gram-negative bacteria. Pseudomonas aeruginosa expresses two T3SSs, namely, an injectisome (iT3SS) translocating effector proteins in the host cell cytosol and a flagellum (fT3SS) ensuring bacterial motility. Inhibiting these systems is an appealing therapeutic strategy for acute infections. This study examines the protective effects of the salicylidene acylhydrazide INP0341 and of the hydroxyquinoline INP1750 (previously described as T3SS inhibitors in other species) toward cytotoxic effects of P. aeruginosain vitro Both compounds reduced cell necrosis and inflammasome activation induced by reference strains or clinical isolates expressing T3SS toxins or only the translocation apparatus. INP0341 inhibited iT3SS transcriptional activation, including in strains with constitutive iT3SS expression, and reduced the total expression of toxins, suggesting it targets iT3SS gene transcription. INP1750 inhibited toxin secretion and flagellar motility and impaired the activity of the YscN ATPase from Yersinia pseudotuberculosis (homologous to the ATPase present in the basal body of P. aeruginosa iT3SS and fT3SS), suggesting that it rather targets a T3SS core constituent with high homology among iT3SS and fT3SS. This mode of action is similar to that previously described for INP1855, another hydroxyquinoline, against P. aeruginosa Thus, although acting by different mechanisms, INP0341 and INP1750 appear as useful inhibitors of the virulence of P. aeruginosa Hydroxyquinolines may have a broader spectrum of activity by the fact they act upon two virulence factors (iT3SS and fT3SS).
Keywords: NLRC4 inflammasome; Pseudomonas aeruginosa; flagella; inhibitors; type three secretion system; virulence.
Copyright © 2017 American Society for Microbiology.
Figures
References
-
- Pena C, Cabot G, Gomez-Zorrilla S, Zamorano L, Ocampo-Sosa A, Murillas J, Almirante B, Pomar V, Aguilar M, Granados A, Calbo E, Rodriguez-Bano J, Rodriguez-Lopez F, Tubau F, Martinez-Martinez L, Oliver A. 2014. Influence of virulence genotype and resistance profile in the mortality of Pseudomonas aeruginosa bloodstream infections. Clin Infect Dis 60:539–548. doi: 10.1093/cid/ciu866. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
