

Synapses in the spotlight with synthetic optogenetics
- PMID: 28396573
- PMCID: PMC5412910
- DOI: 10.15252/embr.201744010
Synapses in the spotlight with synthetic optogenetics
Abstract
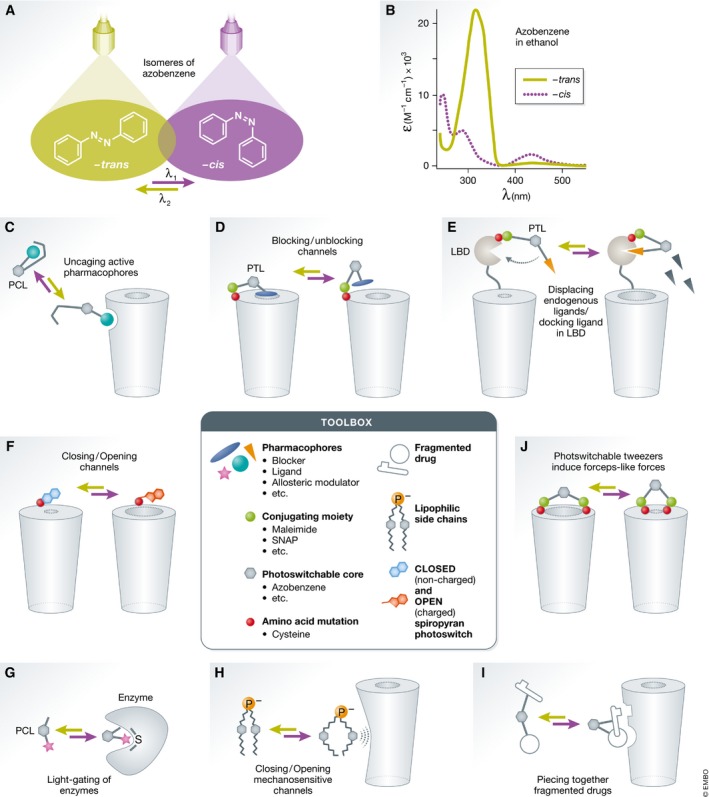
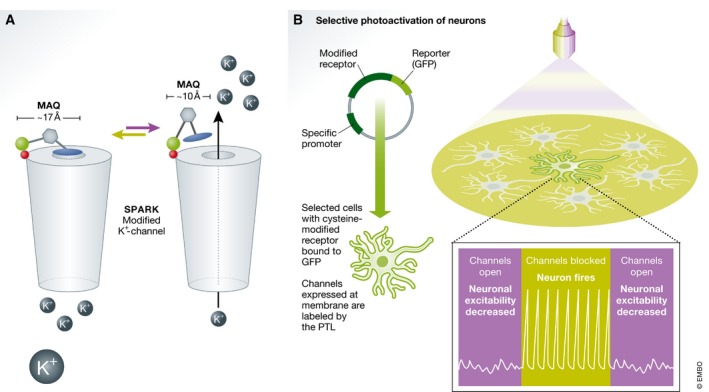
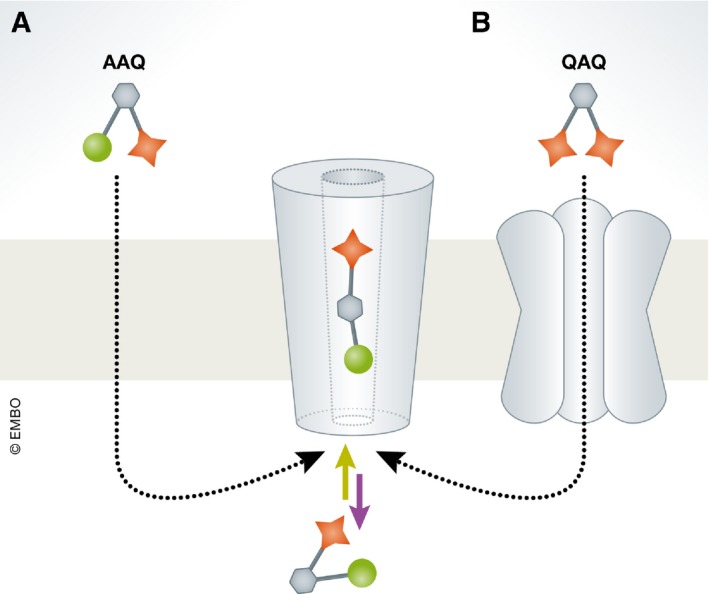
Membrane receptors and ion channels respond to various stimuli and relay that information across the plasma membrane by triggering specific and timed processes. These include activation of second messengers, allowing ion permeation, and changing cellular excitability, to name a few. Gaining control over equivalent processes is essential to understand neuronal physiology and pathophysiology. Recently, new optical techniques have emerged proffering new remote means to control various functions of defined neuronal populations by light, dubbed optogenetics. Still, optogenetic tools do not typically address the activity of receptors and channels native to neurons (or of neuronal origin), nor gain access to their signaling mechanisms. A related method-synthetic optogenetics-bridges this gap by endowing light sensitivity to endogenous neuronal receptors and channels by the appending of synthetic, light-receptive molecules, or photoswitches. This provides the means to photoregulate neuronal receptors and channels and tap into their native signaling mechanisms in select regions of the neurons, such as the synapse. This review discusses the development of synthetic optogenetics as a means to study neuronal receptors and channels remotely, in their natural environment, with unprecedented spatial and temporal precision, and provides an overview of tool design, mode of action, potential clinical applications and insights and achievements gained.
Keywords: native receptors; neurons; optogenetics; photoswitches; synthetic optogenetics.
© 2017 The Authors.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
