

A novel action of follicle-stimulating hormone in the ovary promotes estradiol production without inducing excessive follicular growth before puberty
- PMID: 28397811
- PMCID: PMC5387682
- DOI: 10.1038/srep46222
A novel action of follicle-stimulating hormone in the ovary promotes estradiol production without inducing excessive follicular growth before puberty
Abstract
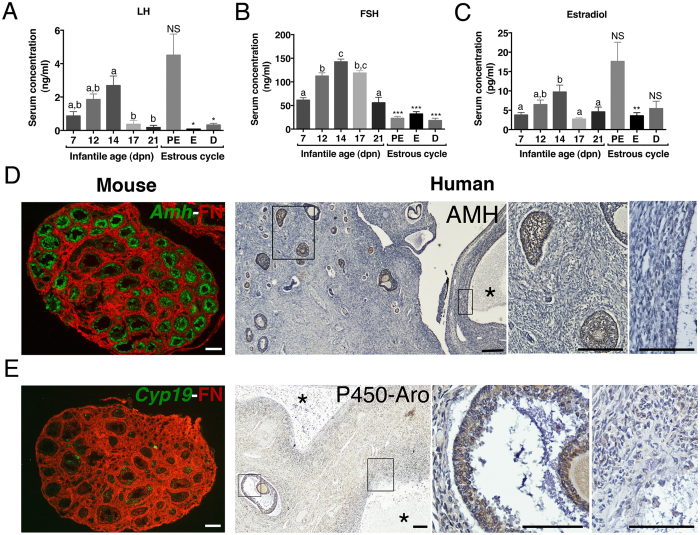
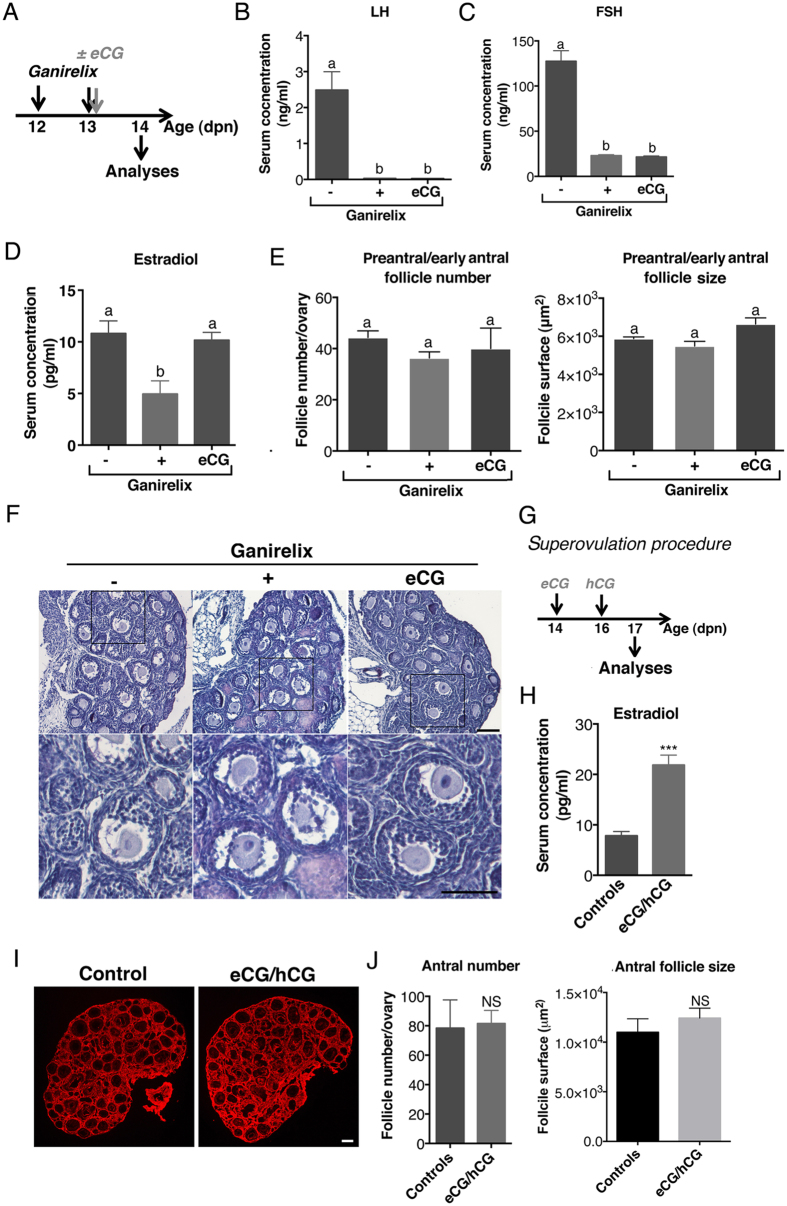
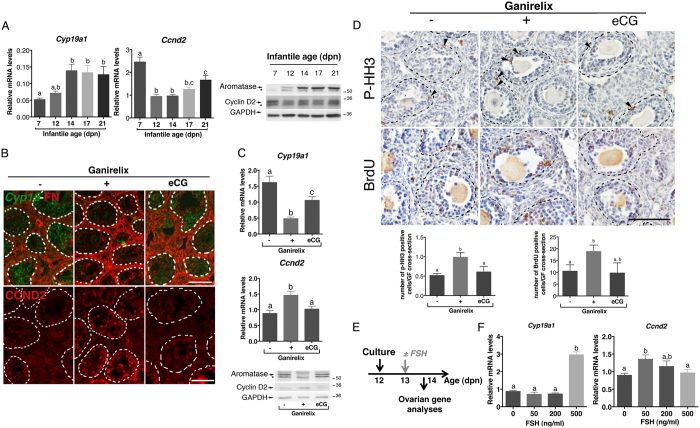
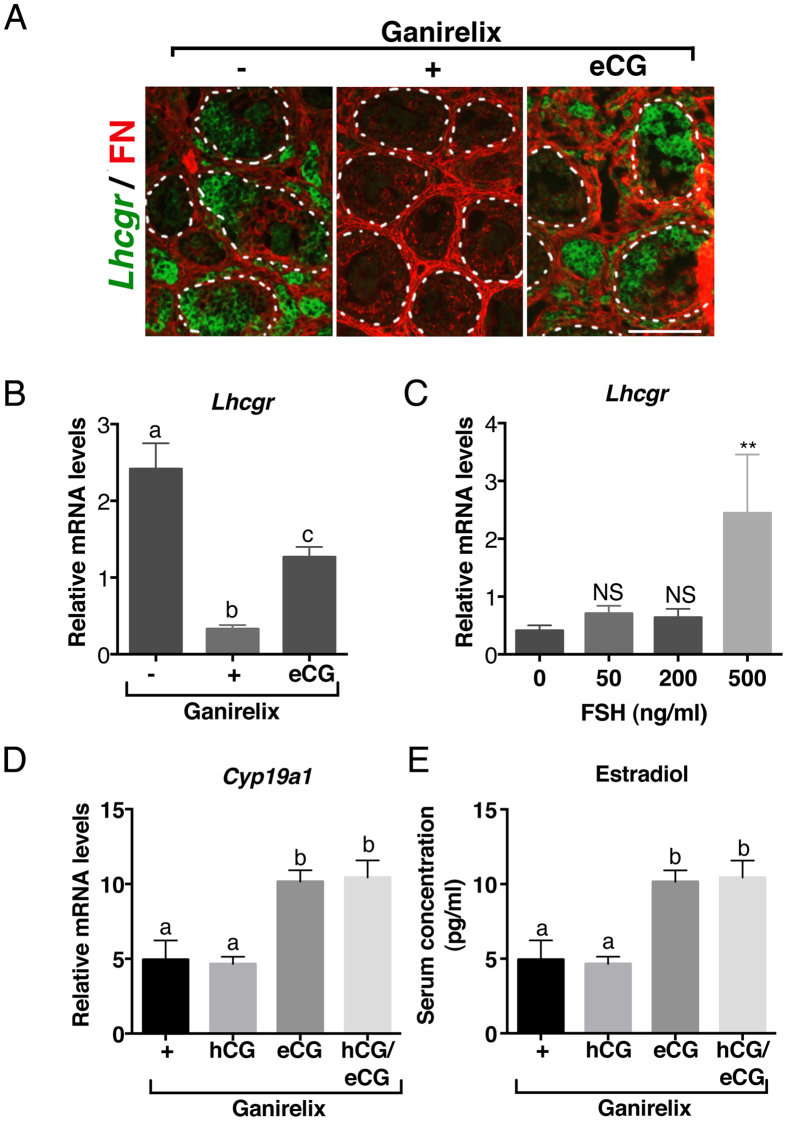
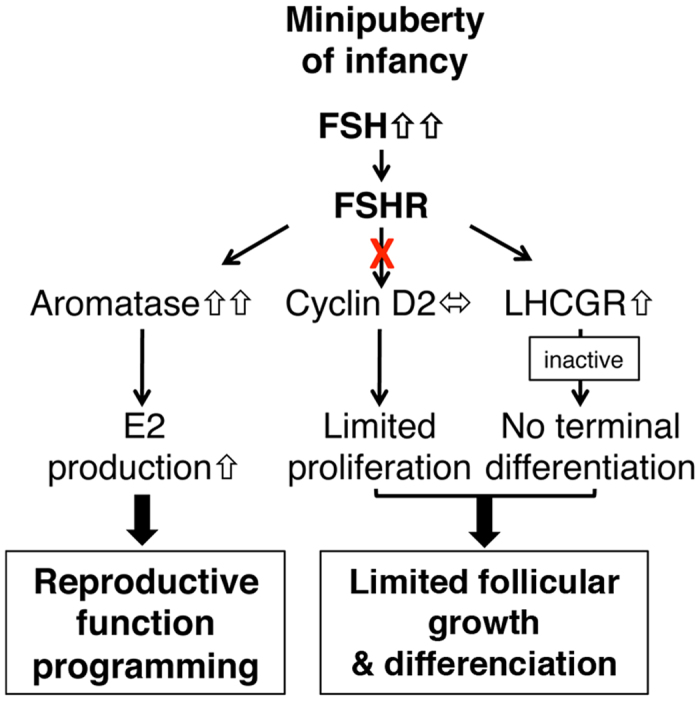
In cyclic females, FSH stimulates ovarian estradiol (E2) production and follicular growth up to the terminal stage. A transient elevation in circulating FSH and E2 levels occurs shortly after birth. But what could be the action of FSH on the ovary during this period, and in particular how it stimulates ovarian steroidogenesis without supporting terminal follicular maturation is intriguing. By experimentally manipulating FSH levels, we demonstrate in mice that the mid-infantile elevation in FSH is mandatory for E2 production by the immature ovary, but that it does not stimulate follicle growth. Importantly, FSH increases aromatase expression to stimulate E2 synthesis, however it becomes unable to induce cyclin D2, a major driver of granulosa cell proliferation. Besides, although FSH prematurely induces luteinizing hormone (LH) receptor expression in granulosa cells, LH pathway is not functional in these cells to induce their terminal differentiation. In line with these results, supplying infantile mice with a superovulation regimen exacerbates E2 production, but it does not stimulate the growth of follicles and it does not induce ovulation. Overall, our findings unveil a regulation whereby high postnatal FSH concentrations ensure the supply of E2 required for programming adult reproductive function without inducing follicular maturation before puberty.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
FSH inhibits AMH to support ovarian estradiol synthesis in infantile mice.J Endocrinol. 2019 Feb 1;240(2):215-228. doi: 10.1530/JOE-18-0313. J Endocrinol. 2019. PMID: 30403655
-
Hypophysectomy of the cyclic mouse. II. Effects of follicle-stimulating hormone (FSH) and luteinizing hormone on folliculogenesis, FSH and human chorionic gonadotropin receptors, and steroidogenesis.Biol Reprod. 1993 Mar;48(3):595-605. doi: 10.1095/biolreprod48.3.595. Biol Reprod. 1993. PMID: 8452936
-
Regulation of ovarian steroidogenesis in vitro by follicle-stimulating hormone and luteinizing hormone during sexual maturation in salmonid fish.Biol Reprod. 2000 May;62(5):1262-9. doi: 10.1095/biolreprod62.5.1262. Biol Reprod. 2000. PMID: 10775175
-
Ovarian follicular and luteal physiology.Int Rev Physiol. 1980;22:117-201. Int Rev Physiol. 1980. PMID: 6248477 Review.
-
Inhibin-B secretion and FSH isoform distribution may play an integral part of follicular selection in the natural menstrual cycle.Mol Hum Reprod. 2017 Jan;23(1):16-24. doi: 10.1093/molehr/gaw070. Epub 2016 Oct 18. Mol Hum Reprod. 2017. PMID: 27756855 Review.
Cited by
-
Androgen receptor signaling regulates follicular growth and steroidogenesis in interaction with gonadotropins in the ovary during mini-puberty in mice.Front Endocrinol (Lausanne). 2023 Apr 19;14:1130681. doi: 10.3389/fendo.2023.1130681. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 37152943 Free PMC article.
-
Assessing the Ability of Developmentally Precocious Estrogen Signaling to Recapitulate Ovarian Transcriptomes and Follicle Dynamics in Alligators from a Contaminated Lake.Environ Health Perspect. 2020 Nov;128(11):117003. doi: 10.1289/EHP6627. Epub 2020 Nov 13. Environ Health Perspect. 2020. PMID: 33186072 Free PMC article.
-
Effects of minipuberty disruption on the expression of sexual behavior in female mice.Sci Rep. 2024 Dec 28;14(1):31297. doi: 10.1038/s41598-024-82653-8. Sci Rep. 2024. PMID: 39732805 Free PMC article.
-
The impact of non-caloric artificial sweetener aspartame on female reproductive system in mice model.Reprod Biol Endocrinol. 2023 Aug 14;21(1):73. doi: 10.1186/s12958-023-01115-4. Reprod Biol Endocrinol. 2023. PMID: 37580716 Free PMC article.
-
Deciphering the Roles & Regulation of Estradiol Signaling during Female Mini-Puberty: Insights from Mouse Models.Int J Mol Sci. 2022 Nov 8;23(22):13695. doi: 10.3390/ijms232213695. Int J Mol Sci. 2022. PMID: 36430167 Free PMC article. Review.
References
-
- Döhler K. D. & Wuttke W. Changes with age in levels of serum gonadotropins, prolactin and gonadal steroids in prepubertal male and female rats. Endocrinology 97, 898–907 (1975). - PubMed
-
- Winter J. S., Faiman C., Hobson W. C., Prasad A. V. & Reyes F. I. Pituitary-gonadal relations in infancy. I. Patterns of serum gonadotropin concentrations from birth to four years of age in man and chimpanzee. J. Clin. Endocrinol. Metab. 40, 545–551 (1975). - PubMed
-
- Dullaart J., Kent J. & Ryle M. Serum gonadotrophin concentrations in infantile female mice. J. Reprod. Fertil. 43, 189–192 (1975). - PubMed
-
- Chellakooty M. et al.. Inhibin A, inhibin B, follicle-stimulating hormone, luteinizing hormone, estradiol, and sex hormone-binding globulin levels in 473 healthy infant girls. J. Clin. Endocrinol. Metab. 88, 3515–3520 (2003). - PubMed
-
- Kuiri-Hänninen T., Sankilampi U. & Dunkel L. Activation of the hypothalamic-pituitary-gonadal axis in infancy: minipuberty. Horm. Res. Pædiatrics 82, 73–80 (2014). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
