

Control of gdhR Expression in Neisseria gonorrhoeae via Autoregulation and a Master Repressor (MtrR) of a Drug Efflux Pump Operon
- PMID: 28400529
- PMCID: PMC5388806
- DOI: 10.1128/mBio.00449-17
Control of gdhR Expression in Neisseria gonorrhoeae via Autoregulation and a Master Repressor (MtrR) of a Drug Efflux Pump Operon
Abstract
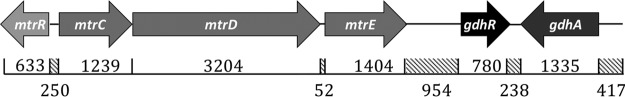
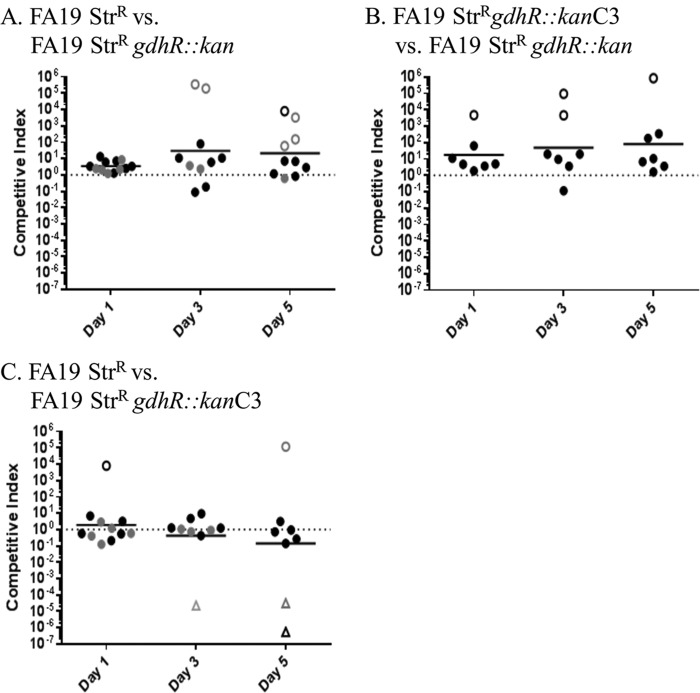
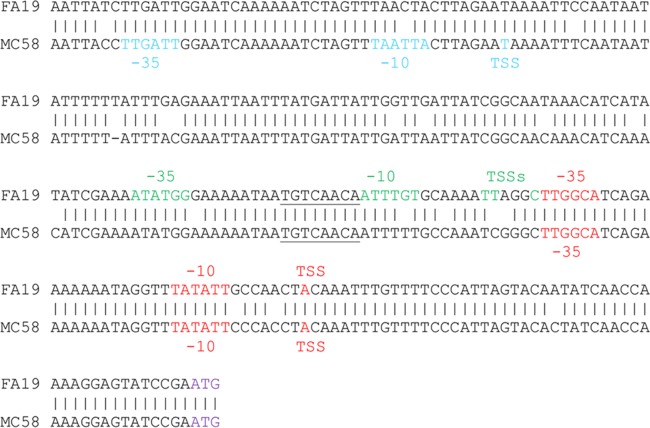
The MtrCDE efflux pump of Neisseria gonorrhoeae contributes to gonococcal resistance to a number of antibiotics used previously or currently in treatment of gonorrhea, as well as to host-derived antimicrobials that participate in innate defense. Overexpression of the MtrCDE efflux pump increases gonococcal survival and fitness during experimental lower genital tract infection of female mice. Transcription of mtrCDE can be repressed by the DNA-binding protein MtrR, which also acts as a global regulator of genes involved in important metabolic, physiologic, or regulatory processes. Here, we investigated whether a gene downstream of mtrCDE, previously annotated gdhR in Neisseria meningitidis, is a target for regulation by MtrR. In meningococci, GdhR serves as a regulator of genes involved in glucose catabolism, amino acid transport, and biosynthesis, including gdhA, which encodes an l-glutamate dehydrogenase and is located next to gdhR but is transcriptionally divergent. We report here that in N. gonorrhoeae, expression of gdhR is subject to autoregulation by GdhR and direct repression by MtrR. Importantly, loss of GdhR significantly increased gonococcal fitness compared to a complemented mutant strain during experimental murine infection. Interestingly, loss of GdhR did not influence expression of gdhA, as reported for meningococci. This variance is most likely due to differences in promoter localization and utilization between gonococci and meningococci. We propose that transcriptional control of gonococcal genes through the action of MtrR and GdhR contributes to fitness of N. gonorrhoeae during infection.IMPORTANCE The pathogenic Neisseria species are strict human pathogens that can cause a sexually transmitted infection (N. gonorrhoeae) or meningitis or fulminant septicemia (N. meningitidis). Although they share considerable genetic information, little attention has been directed to comparing transcriptional regulatory systems that modulate expression of their conserved genes. We hypothesized that transcriptional regulatory differences exist between these two pathogens, and we used the gdh locus as a model to test this idea. For this purpose, we studied two conserved genes (gdhR and gdhA) within the locus. Despite general conservation of the gdh locus in gonococci and meningococci, differences exist in noncoding sequences that correspond to promoter elements or potential sites for interacting with DNA-binding proteins, such as GdhR and MtrR. Our results indicate that implications drawn from studying regulation of conserved genes in one pathogen are not necessarily translatable to a genetically related pathogen.
Keywords: efflux pumps; gonococci; physiology; transcription.
Copyright © 2017 Rouquette-Loughlin et al.
Figures



Similar articles
-
Gonococcal Clinical Strains Bearing a Common gdhR Single Nucleotide Polymorphism That Results in Enhanced Expression of the Virulence Gene lctP Frequently Possess a mtrR Promoter Mutation That Decreases Antibiotic Susceptibility.mBio. 2022 Apr 26;13(2):e0027622. doi: 10.1128/mbio.00276-22. Epub 2022 Mar 8. mBio. 2022. PMID: 35258329 Free PMC article.
-
Could Dampening Expression of the Neisseria gonorrhoeae mtrCDE-Encoded Efflux Pump Be a Strategy To Preserve Currently or Resurrect Formerly Used Antibiotics To Treat Gonorrhea?mBio. 2019 Aug 13;10(4):e01576-19. doi: 10.1128/mBio.01576-19. mBio. 2019. PMID: 31409679 Free PMC article.
-
Dueling regulatory properties of a transcriptional activator (MtrA) and repressor (MtrR) that control efflux pump gene expression in Neisseria gonorrhoeae.mBio. 2012 Dec 4;3(6):e00446-12. doi: 10.1128/mBio.00446-12. mBio. 2012. PMID: 23221802 Free PMC article.
-
Transcriptional regulation of the mtrCDE efflux pump operon: importance for Neisseria gonorrhoeae antimicrobial resistance.Microbiology (Reading). 2022 Aug;168(8):001231. doi: 10.1099/mic.0.001231. Microbiology (Reading). 2022. PMID: 35916832 Free PMC article. Review.
-
Genetic organization and regulation of antimicrobial efflux systems possessed by Neisseria gonorrhoeae and Neisseria meningitidis.J Mol Microbiol Biotechnol. 2001 Apr;3(2):219-24. J Mol Microbiol Biotechnol. 2001. PMID: 11321577 Review.
Cited by
-
Transcriptional regulation of a gonococcal gene encoding a virulence factor (L-lactate permease).PLoS Pathog. 2019 Dec 20;15(12):e1008233. doi: 10.1371/journal.ppat.1008233. eCollection 2019 Dec. PLoS Pathog. 2019. PMID: 31860664 Free PMC article.
-
Structures of Neisseria gonorrhoeae MtrR-operator complexes reveal molecular mechanisms of DNA recognition and antibiotic resistance-conferring clinical mutations.Nucleic Acids Res. 2021 Apr 19;49(7):4155-4170. doi: 10.1093/nar/gkab213. Nucleic Acids Res. 2021. PMID: 33784401 Free PMC article.
-
Mechanistic Basis for Decreased Antimicrobial Susceptibility in a Clinical Isolate of Neisseria gonorrhoeae Possessing a Mosaic-Like mtr Efflux Pump Locus.mBio. 2018 Nov 27;9(6):e02281-18. doi: 10.1128/mBio.02281-18. mBio. 2018. PMID: 30482834 Free PMC article.
-
Neisseria meningitidis Sibling Small Regulatory RNAs Connect Metabolism with Colonization by Controlling Propionate Use.J Bacteriol. 2023 Mar 21;205(3):e0046222. doi: 10.1128/jb.00462-22. Epub 2023 Mar 1. J Bacteriol. 2023. PMID: 36856428 Free PMC article.
-
Gonococcal Clinical Strains Bearing a Common gdhR Single Nucleotide Polymorphism That Results in Enhanced Expression of the Virulence Gene lctP Frequently Possess a mtrR Promoter Mutation That Decreases Antibiotic Susceptibility.mBio. 2022 Apr 26;13(2):e0027622. doi: 10.1128/mbio.00276-22. Epub 2022 Mar 8. mBio. 2022. PMID: 35258329 Free PMC article.
References
-
- Newman L, Rowley J, Vander Hoorn S, Wijesooriya NS, Unemo M, Low N, Stevens G, Gottlieb S, Kiarie J, Temmerman M. 2015. Global estimates of the prevalence and incidence of four curable sexually transmitted infections in 2012 based on systematic review and global reporting. PLoS One 10:e0143304. doi:10.1371/journal.pone.0143304. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
