

The Periplasmic Chaperone Network of Campylobacter jejuni: Evidence that SalC (Cj1289) and PpiD (Cj0694) Are Involved in Maintaining Outer Membrane Integrity
- PMID: 28400767
- PMCID: PMC5368265
- DOI: 10.3389/fmicb.2017.00531
The Periplasmic Chaperone Network of Campylobacter jejuni: Evidence that SalC (Cj1289) and PpiD (Cj0694) Are Involved in Maintaining Outer Membrane Integrity
Abstract
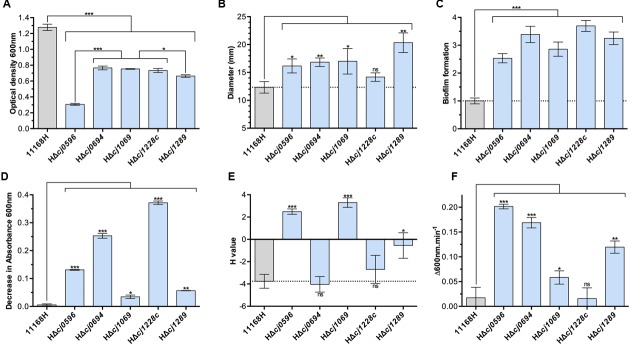
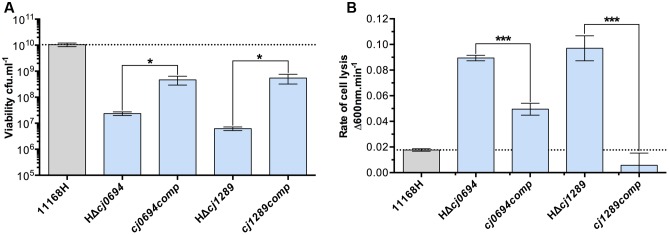
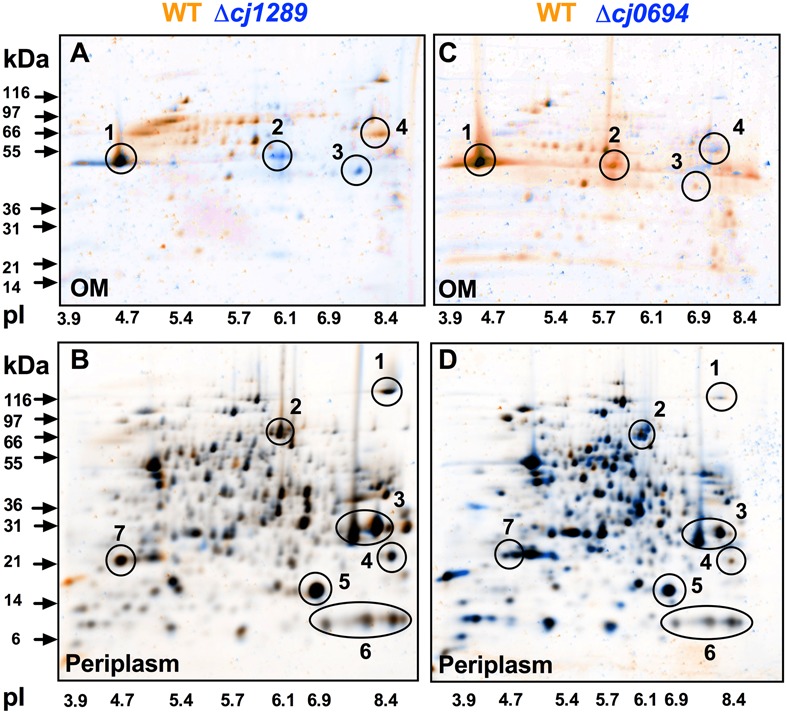
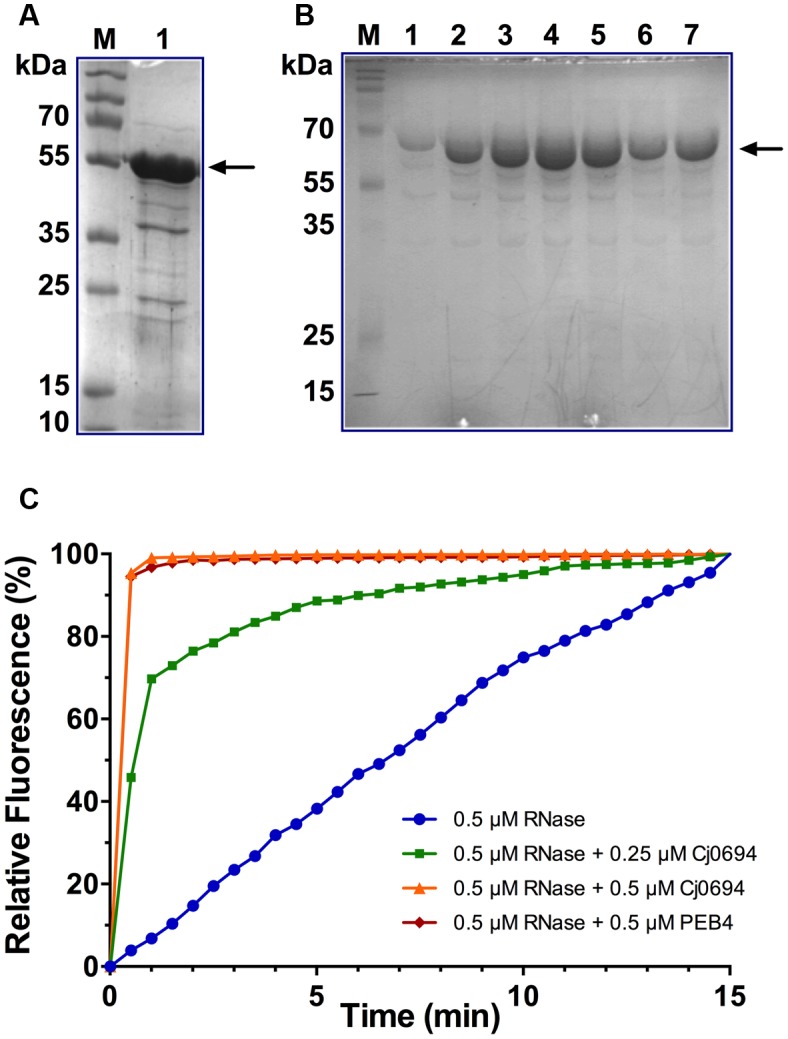
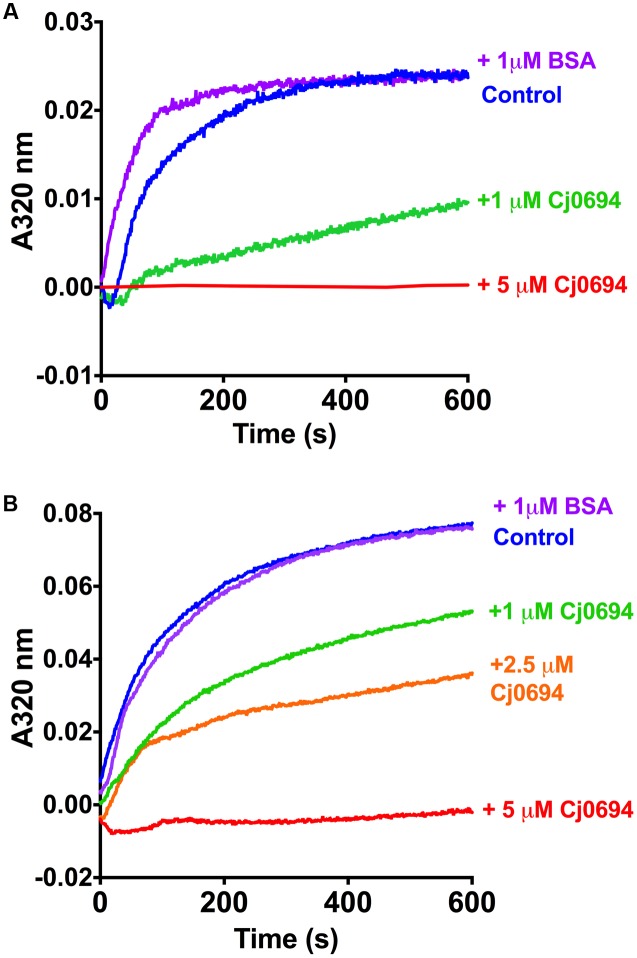
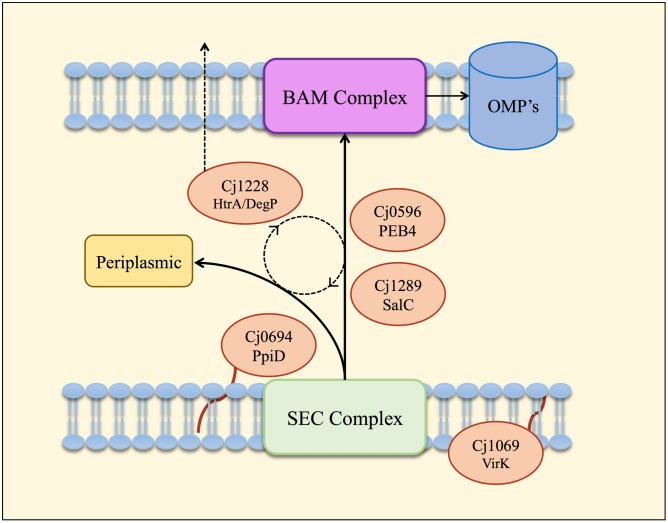
The outer membrane (OM) of Gram-negative pathogenic bacteria is a key structure in host-pathogen interactions that contains a plethora of proteins, performing a range of functions including adhesion, nutrient uptake, export of effectors and interaction with innate and adaptive components of the immune system. In addition, the OM can exclude drugs and thus contribute to antimicrobial resistance. The OM of the food-borne pathogen Campylobacter jejuni contains porins, adhesins and other virulence factors that must be specifically localized to this membrane, but the protein sorting mechanisms involved are only partially understood. In particular, chaperones are required to ferry OM proteins across the periplasm after they emerge from the Sec translocation system. The SurA-related chaperone PEB4 (Cj0596) is the only protein with a proven role in OM biogenesis and integrity in C. jejuni. In this work, we have constructed a set of isogenic deletion mutants in genes encoding both known and predicted chaperones (cj0596, cj0694, cj1069, cj1228c, and cj1289) using NCTC 11168H as the parental strain. These mutants were characterized using a range of assays to determine effects on growth, agglutination, biofilm formation, membrane permeability and hydrophobicity. We focused on Cj1289 and Cj0694, which our previous work suggested possessed both chaperone and peptidyl-proyl cis/trans isomerase (PPIase) domains. Mutants in either cj1289 or cj0694 showed growth defects, increased motility, agglutination and biofilm formation and severe OM permeability defects as measured by a lysozyme accessibility assay, that were comparable to those exhibited by the isogenic peb4 mutant. 2D-gel comparisons showed a general decrease in OM proteins in these mutants. We heterologously overproduced and purified Cj0694 and obtained evidence that this protein was an active PPIase, as judged by its acceleration of the refolding rate of reduced and alkylated ribonuclease T1 and that it also possessed holdase-type chaperone activity. Cj0694 is most similar to the PpiD class of chaperones but is unusual in possessing PPIase activity. Taken together, our data show that in addition to PEB4, Cj1289 (SalC; SurA-like chaperone) and Cj0694 (PpiD) are also key proteins involved in OM biogenesis and integrity in C. jejuni.
Keywords: Campylobacter; HtrA; PEB4; PpiD; SurA; VirK; outer membrane; periplasmic chaperone.
Figures
Similar articles
-
The virulence factor PEB4 (Cj0596) and the periplasmic protein Cj1289 are two structurally related SurA-like chaperones in the human pathogen Campylobacter jejuni.J Biol Chem. 2011 Jun 17;286(24):21254-65. doi: 10.1074/jbc.M111.220442. Epub 2011 Apr 26. J Biol Chem. 2011. PMID: 21524997 Free PMC article.
-
PpiD is a player in the network of periplasmic chaperones in Escherichia coli.BMC Microbiol. 2010 Sep 29;10:251. doi: 10.1186/1471-2180-10-251. BMC Microbiol. 2010. PMID: 20920237 Free PMC article.
-
Conformational dynamics of Peb4 exhibit "mother's arms" chain model: a molecular dynamics study.J Biomol Struct Dyn. 2017 Aug;35(10):2186-2196. doi: 10.1080/07391102.2016.1209131. Epub 2016 Jul 19. J Biomol Struct Dyn. 2017. PMID: 27434141
-
The role of SurA factor in outer membrane protein transport and virulence.Int J Med Microbiol. 2010 Nov;300(7):421-8. doi: 10.1016/j.ijmm.2010.04.012. Epub 2010 May 6. Int J Med Microbiol. 2010. PMID: 20447864 Review.
-
The Periplasmic Chaperones Skp and SurA.Subcell Biochem. 2019;92:169-186. doi: 10.1007/978-3-030-18768-2_6. Subcell Biochem. 2019. PMID: 31214987 Review.
Cited by
-
Review on Stress Tolerance in Campylobacter jejuni.Front Cell Infect Microbiol. 2021 Feb 4;10:596570. doi: 10.3389/fcimb.2020.596570. eCollection 2020. Front Cell Infect Microbiol. 2021. PMID: 33614524 Free PMC article. Review.
-
Quantitative Analyses Reveal Novel Roles for N-Glycosylation in a Major Enteric Bacterial Pathogen.mBio. 2019 Apr 23;10(2):e00297-19. doi: 10.1128/mBio.00297-19. mBio. 2019. PMID: 31015322 Free PMC article.
-
Flagellar stator genes control a trophic shift from obligate to facultative predation and biofilm formation in a bacterial predator.mBio. 2024 Aug 14;15(8):e0071524. doi: 10.1128/mbio.00715-24. Epub 2024 Jul 22. mBio. 2024. PMID: 39037271 Free PMC article.
-
The Periplasmic Oxidoreductase DsbA Is Required for Virulence of the Phytopathogen Dickeya solani.Int J Mol Sci. 2022 Jan 9;23(2):697. doi: 10.3390/ijms23020697. Int J Mol Sci. 2022. PMID: 35054882 Free PMC article.
-
Agricultural intensification and the evolution of host specialism in the enteric pathogen Campylobacter jejuni.Proc Natl Acad Sci U S A. 2020 May 19;117(20):11018-11028. doi: 10.1073/pnas.1917168117. Epub 2020 May 4. Proc Natl Acad Sci U S A. 2020. PMID: 32366649 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases