

Autoantibodies against GPIHBP1 as a Cause of Hypertriglyceridemia
- PMID: 28402248
- PMCID: PMC5555413
- DOI: 10.1056/NEJMoa1611930
Autoantibodies against GPIHBP1 as a Cause of Hypertriglyceridemia
Abstract
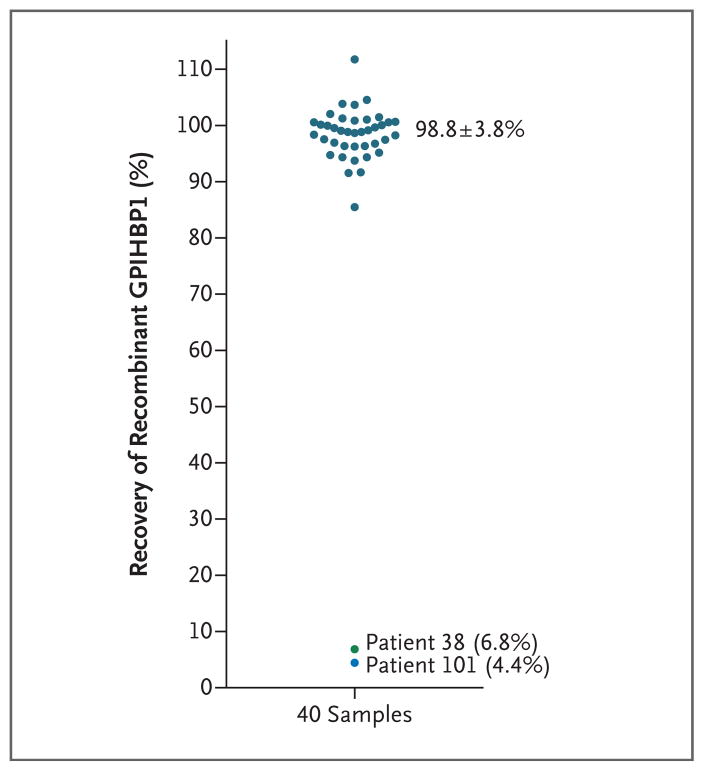
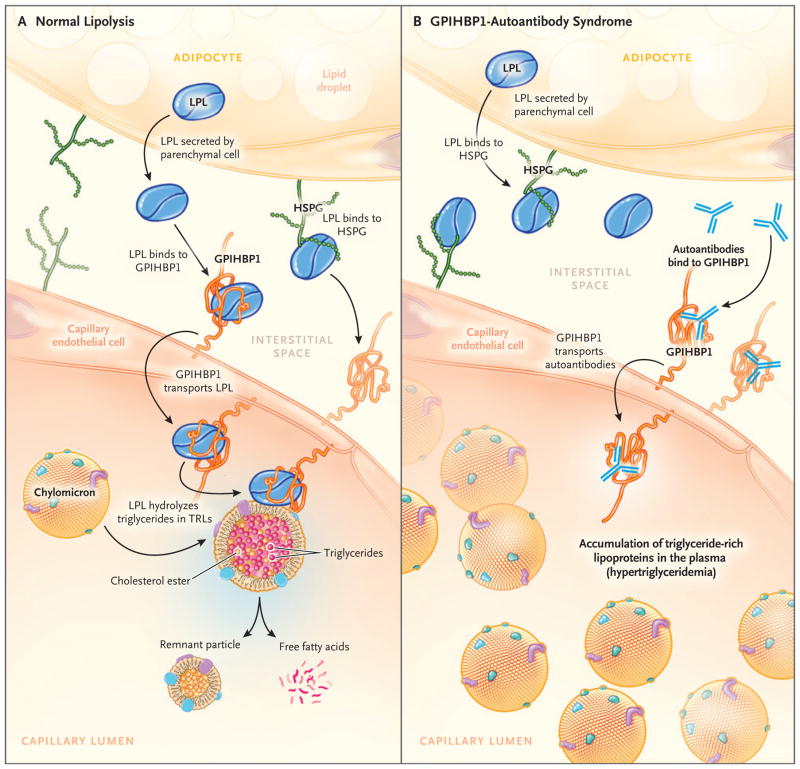
Background: A protein that is expressed on capillary endothelial cells, called GPIHBP1 (glycosylphosphatidylinositol-anchored high-density lipoprotein binding protein 1), binds lipoprotein lipase and shuttles it to its site of action in the capillary lumen. A deficiency in GPIHBP1 prevents lipoprotein lipase from reaching the capillary lumen. Patients with GPIHBP1 deficiency have low plasma levels of lipoprotein lipase, impaired intravascular hydrolysis of triglycerides, and severe hypertriglyceridemia (chylomicronemia). During the characterization of a monoclonal antibody-based immunoassay for GPIHBP1, we encountered two plasma samples (both from patients with chylomicronemia) that contained an interfering substance that made it impossible to measure GPIHBP1. That finding raised the possibility that those samples might contain GPIHBP1 autoantibodies.
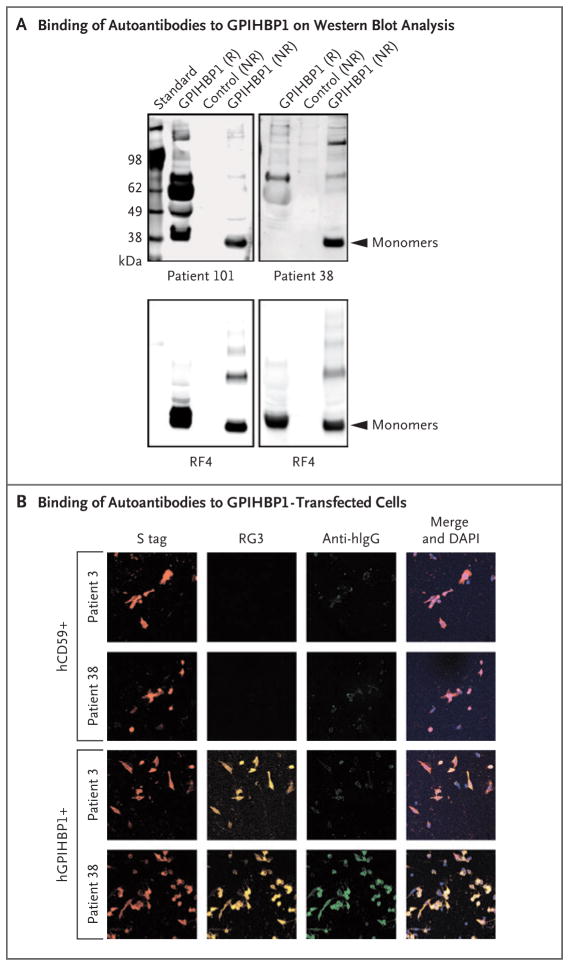
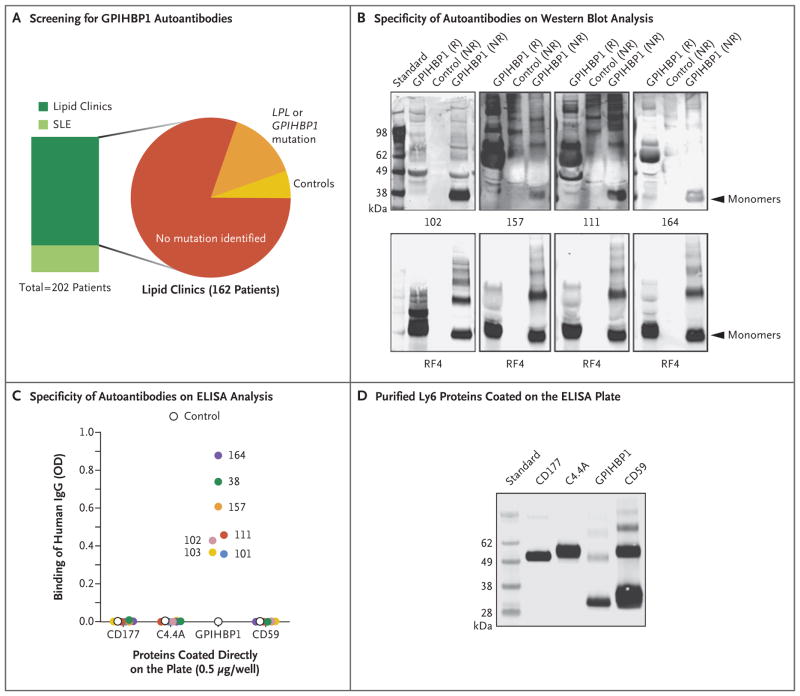
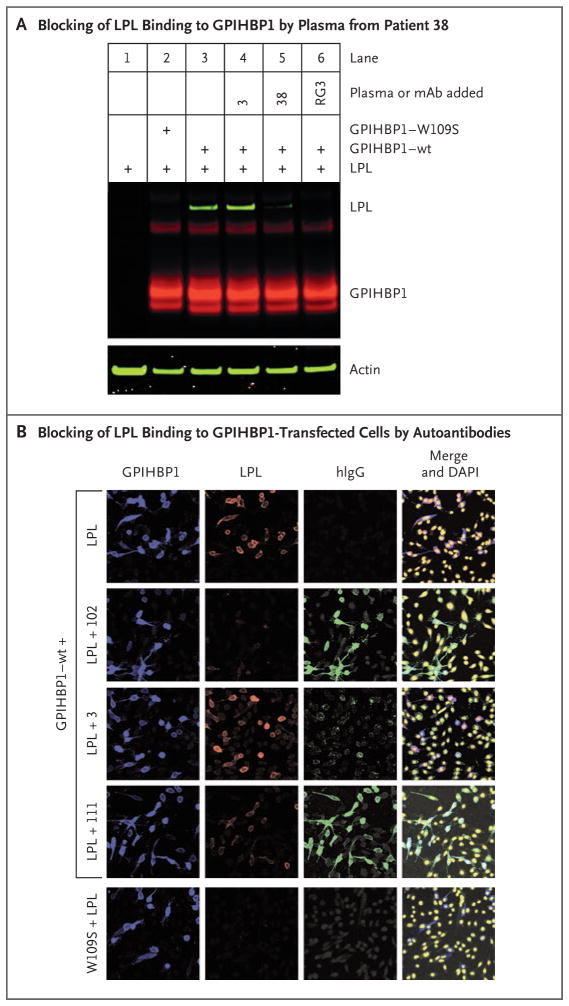
Methods: Using a combination of immunoassays, Western blot analyses, and immunocytochemical studies, we tested the two plasma samples (as well as samples from other patients with chylomicronemia) for the presence of GPIHBP1 autoantibodies. We also tested the ability of GPIHBP1 autoantibodies to block the binding of lipoprotein lipase to GPIHBP1.
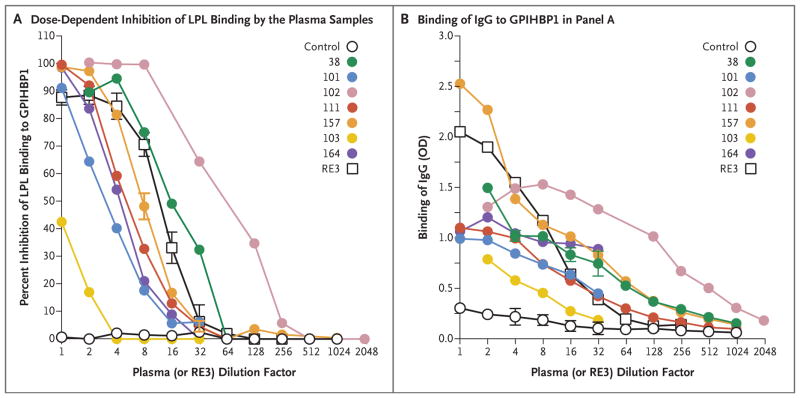
Results: We identified GPIHBP1 autoantibodies in six patients with chylomicronemia and found that these autoantibodies blocked the binding of lipoprotein lipase to GPIHBP1. As in patients with GPIHBP1 deficiency, those with GPIHBP1 autoantibodies had low plasma levels of lipoprotein lipase. Three of the six patients had systemic lupus erythematosus. One of these patients who had GPIHBP1 autoantibodies delivered a baby with plasma containing maternal GPIHBP1 autoantibodies; the infant had severe but transient chylomicronemia. Two of the patients with chylomicronemia and GPIHBP1 autoantibodies had a response to treatment with immunosuppressive agents.
Conclusions: In six patients with chylomicronemia, GPIHBP1 autoantibodies blocked the ability of GPIHBP1 to bind and transport lipoprotein lipase, thereby interfering with lipoprotein lipase-mediated processing of triglyceride-rich lipoproteins and causing severe hypertriglyceridemia. (Funded by the National Heart, Lung, and Blood Institute and the Leducq Foundation.).
Figures
Comment in
-
Dyslipidaemia: Hypertriglyceridaemia caused by GPIHBP1 autoantibodies.Nat Rev Cardiol. 2017 Jun;14(6):316-317. doi: 10.1038/nrcardio.2017.62. Epub 2017 Apr 24. Nat Rev Cardiol. 2017. PMID: 28436485 No abstract available.
-
Triglyceride Metabolism under Attack.Cell Metab. 2017 Jun 6;25(6):1209-1210. doi: 10.1016/j.cmet.2017.05.005. Cell Metab. 2017. PMID: 28591627
References
-
- Charrière S, Peretti N, Bernard S, et al. GPIHBP1 C89F neomutation and hydrophobic C-terminal domain G175R mutation in two pedigrees with severe hyperchylomicronemia. J Clin Endocrinol Metab. 2011;96:E1675–E1679. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials