

Stepwise assembly of the earliest precursors of large ribosomal subunits in yeast
- PMID: 28402444
- PMCID: PMC5499802
- DOI: 10.1093/nar/gkx254
Stepwise assembly of the earliest precursors of large ribosomal subunits in yeast
Abstract
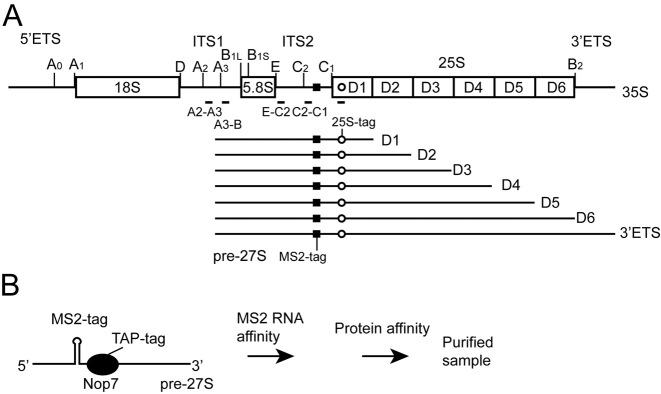
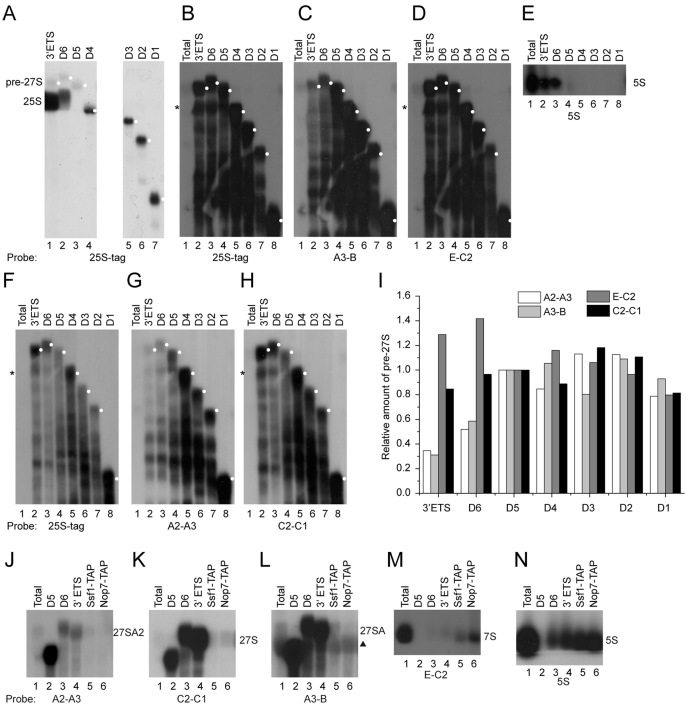
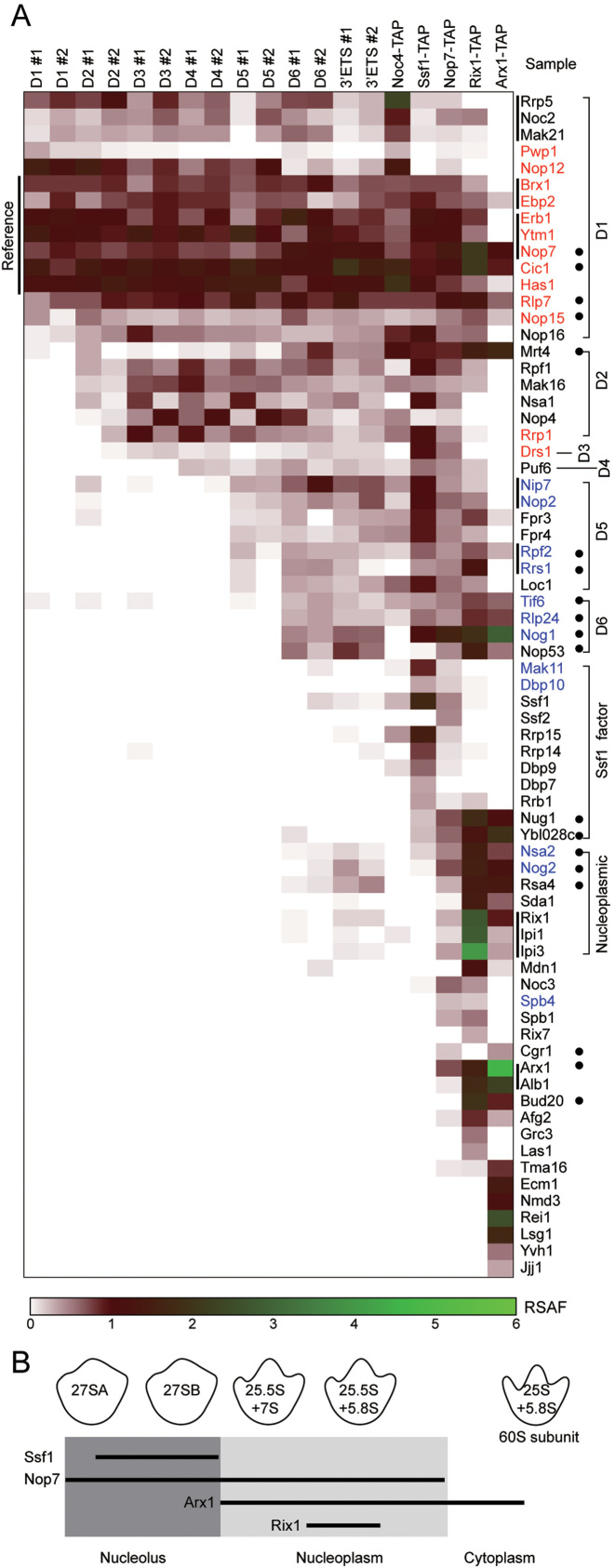
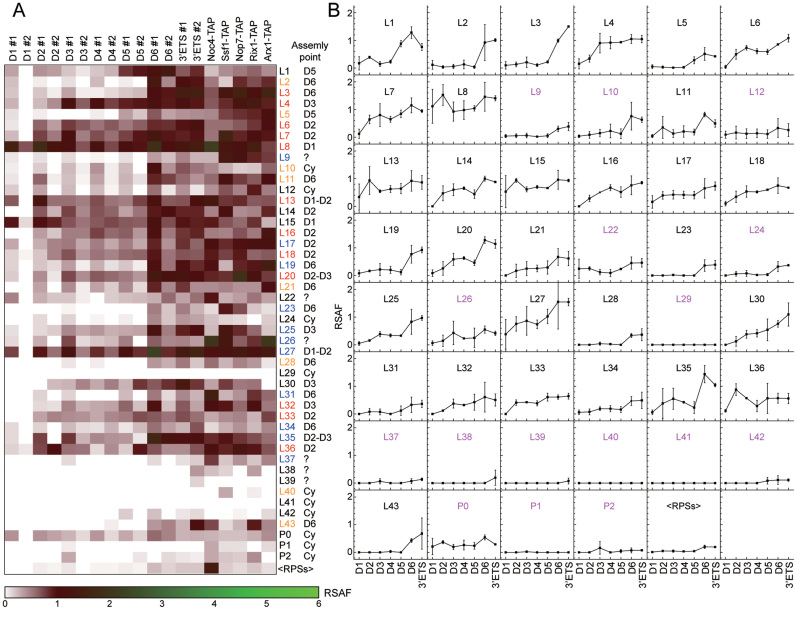
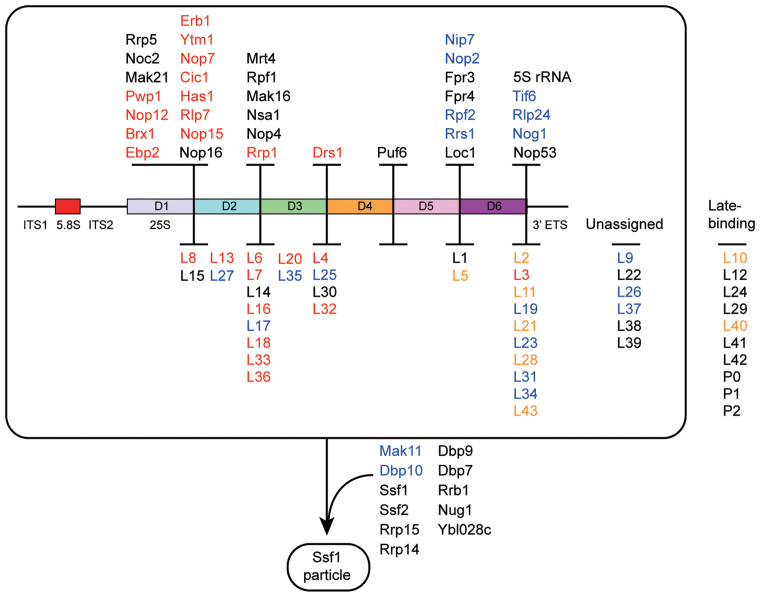
Small ribosomal subunits are co-transcriptionally assembled on the nascent precursor rRNA in Saccharomyces cerevisiae. It is unknown how the highly intertwined structure of 60S large ribosomal subunits is initially formed. Here, we affinity purified and analyzed a series of pre-60S particles assembled in vivo on plasmid-encoded pre-rRNA fragments of increasing lengths, revealing a spatiotemporal assembly map for 34 trans-acting assembly factors (AFs), 30 ribosomal proteins and 5S rRNA. The gradual association of AFs and ribosomal proteins with the pre-rRNA fragments strongly supports that the pre-60S is co-transcriptionally, rather than post-transcriptionally, assembled. The internal and external transcribed spacers ITS1, ITS2 and 3΄ ETS in pre-rRNA must be processed in pre-60S. We show that the processing machineries for ITS1 and ITS2 are primarily recruited by the 5΄ and 3΄ halves of pre-27S RNA, respectively. Nevertheless, processing of both ITS1 and ITS2 requires a complete 25S region. The 3΄ ETS plays a minor role in ribosome assembly, but is important for efficient rRNA processing and ribosome maturation. We also identified a distinct pre-60S state occurring before ITS2 processing. Our data reveal the elusive co-transcriptional assembly pathway of large ribosomal subunit.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Grandi P., Rybin V., Bassler J., Petfalski E., Strauss D., Marzioch M., Schafer T., Kuster B., Tschochner H., Tollervey D. et al. 90S pre-ribosomes include the 35S pre-rRNA, the U3 snoRNP, and 40S subunit processing factors but predominantly lack 60S synthesis factors. Mol. Cell. 2002; 10:105–115. - PubMed
-
- Osheim Y.N., French S.L., Keck K.M., Champion E.A., Spasov K., Dragon F., Baserga S.J., Beyer A.L.. Pre-18S ribosomal RNA is structurally compacted into the SSU processome prior to being cleaved from nascent transcripts in Saccharomyces cerevisiae. Mol. Cell. 2004; 16:943–954. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
