

A novel mechanism of cone photoreceptor adaptation
- PMID: 28403143
- PMCID: PMC5389785
- DOI: 10.1371/journal.pbio.2001210
A novel mechanism of cone photoreceptor adaptation
Abstract
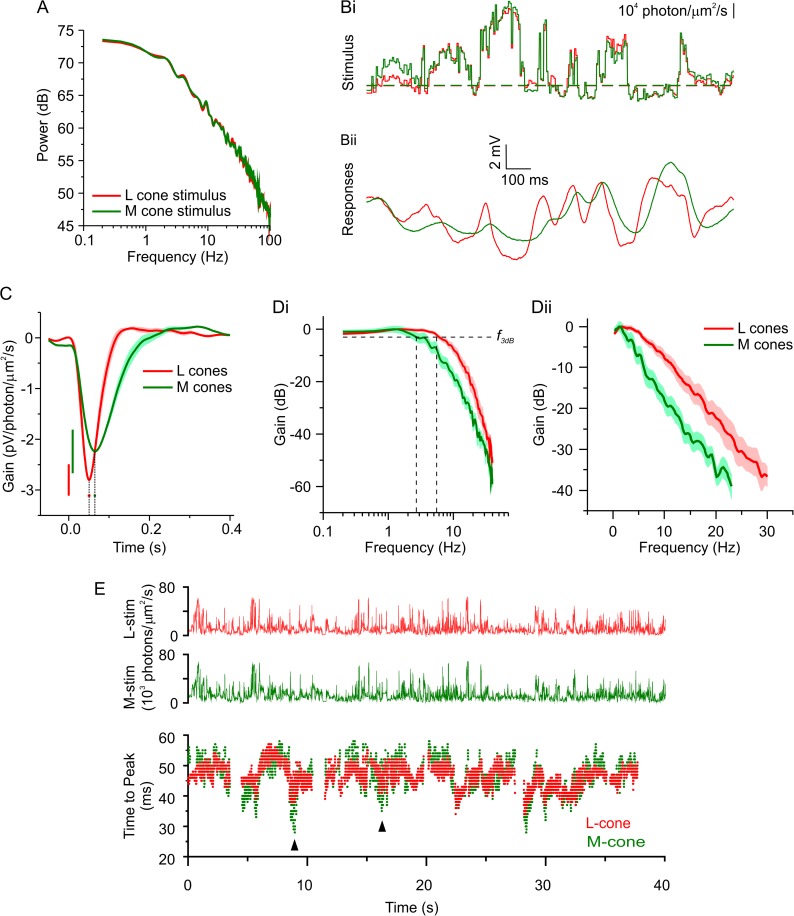
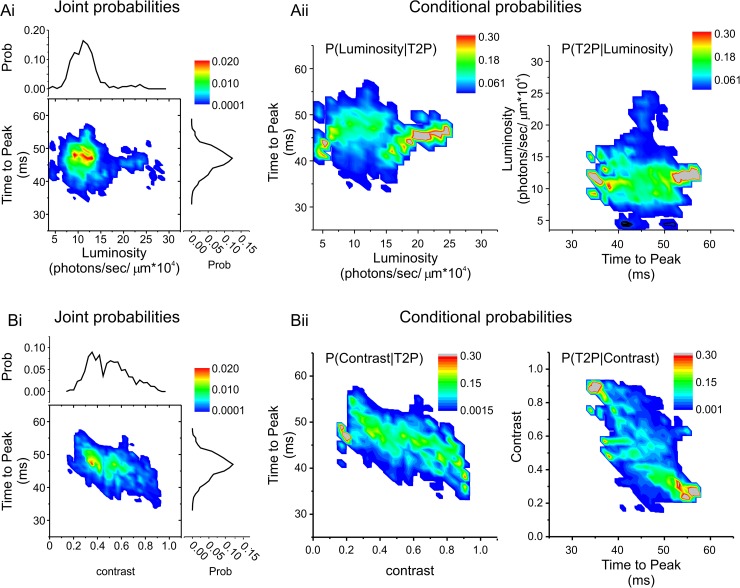
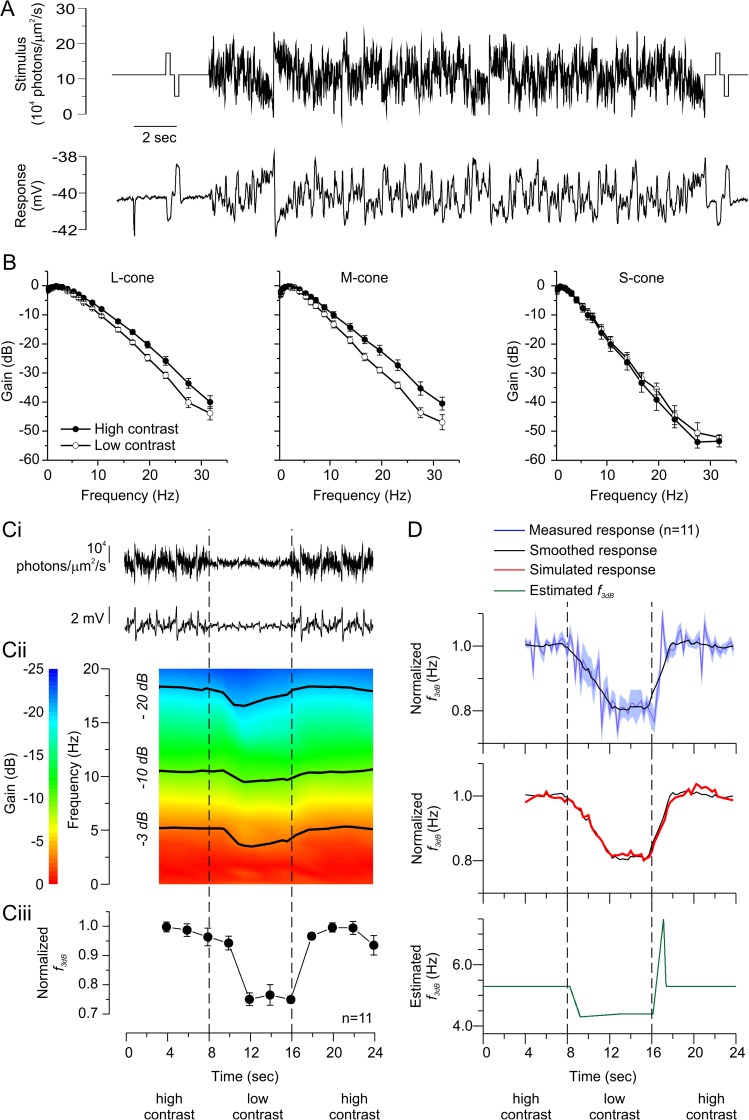
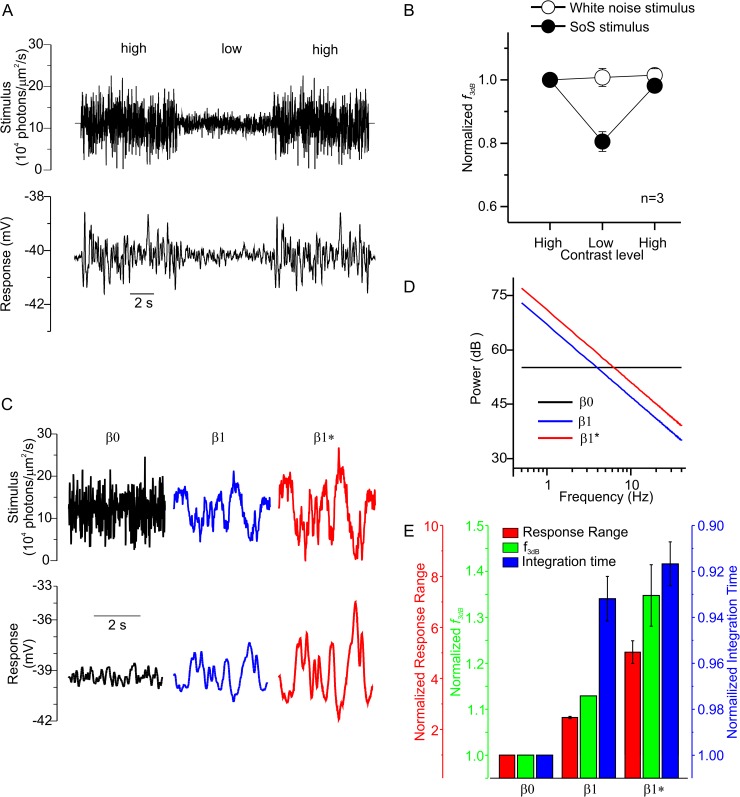
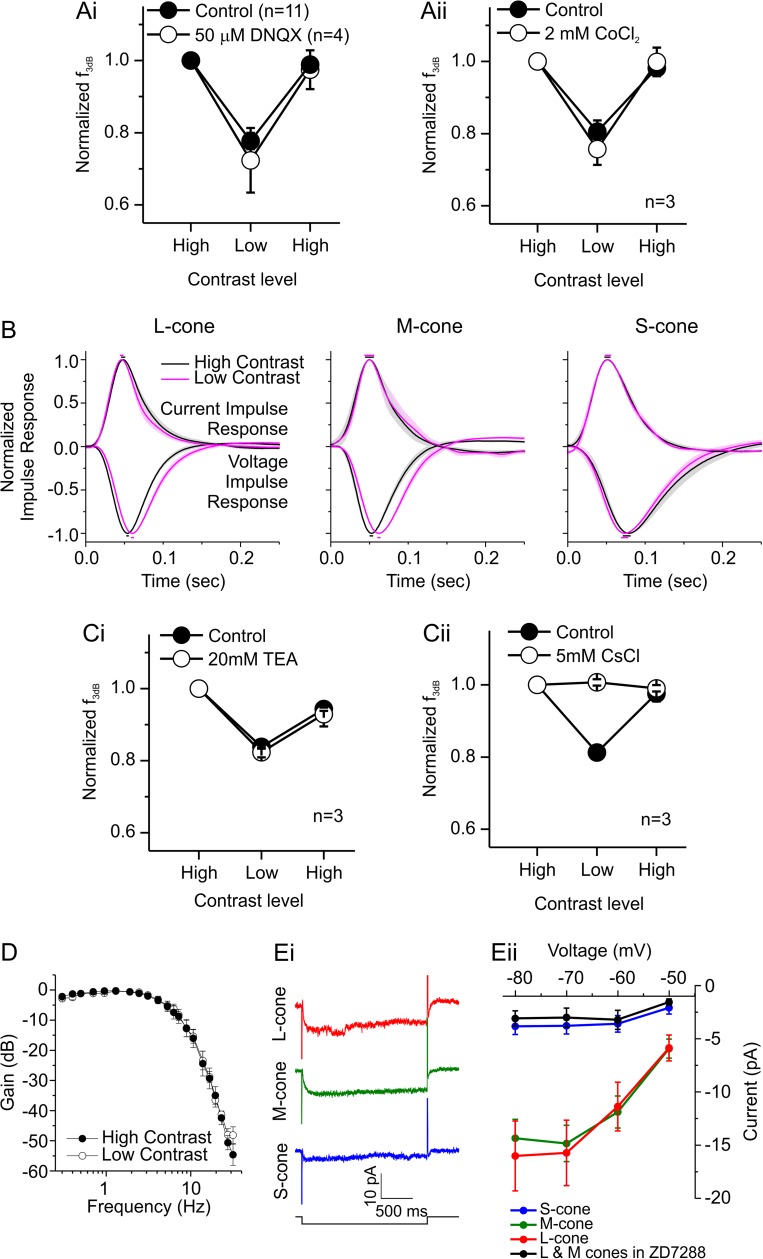
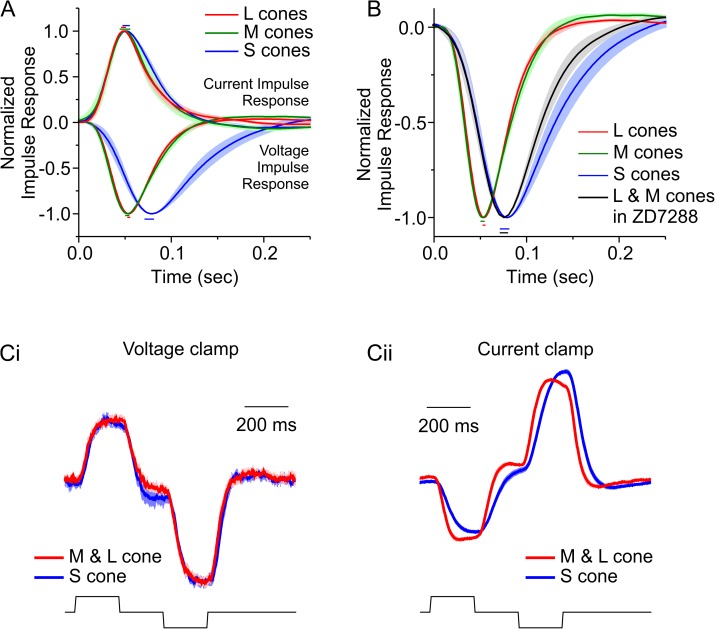
An animal's ability to survive depends on its sensory systems being able to adapt to a wide range of environmental conditions, by maximizing the information extracted and reducing the noise transmitted. The visual system does this by adapting to luminance and contrast. While luminance adaptation can begin at the retinal photoreceptors, contrast adaptation has been shown to start at later stages in the retina. Photoreceptors adapt to changes in luminance over multiple time scales ranging from tens of milliseconds to minutes, with the adaptive changes arising from processes within the phototransduction cascade. Here we show a new form of adaptation in cones that is independent of the phototransduction process. Rather, it is mediated by voltage-gated ion channels in the cone membrane and acts by changing the frequency response of cones such that their responses speed up as the membrane potential modulation depth increases and slow down as the membrane potential modulation depth decreases. This mechanism is effectively activated by high-contrast stimuli dominated by low frequencies such as natural stimuli. However, the more generally used Gaussian white noise stimuli were not effective since they did not modulate the cone membrane potential to the same extent. This new adaptive process had a time constant of less than a second. A critical component of the underlying mechanism is the hyperpolarization-activated current, Ih, as pharmacologically blocking it prevented the long- and mid- wavelength sensitive cone photoreceptors (L- and M-cones) from adapting. Consistent with this, short- wavelength sensitive cone photoreceptors (S-cones) did not show the adaptive response, and we found they also lacked a prominent Ih. The adaptive filtering mechanism identified here improves the information flow by removing higher-frequency noise during lower signal-to-noise ratio conditions, as occurs when contrast levels are low. Although this new adaptive mechanism can be driven by contrast, it is not a contrast adaptation mechanism in its strictest sense, as will be argued in the Discussion.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Regional tuning of photoreceptor adaptation in the primate retina.Nat Commun. 2024 Oct 12;15(1):8821. doi: 10.1038/s41467-024-53061-3. Nat Commun. 2024. PMID: 39394185 Free PMC article.
-
In intact mammalian photoreceptors, Ca2+-dependent modulation of cGMP-gated ion channels is detectable in cones but not in rods.J Gen Physiol. 2004 Jan;123(1):63-75. doi: 10.1085/jgp.200308952. J Gen Physiol. 2004. PMID: 14699078 Free PMC article.
-
Intrinsic cone adaptation modulates feedback efficiency from horizontal cells to cones.J Gen Physiol. 1999 Oct;114(4):511-24. doi: 10.1085/jgp.114.4.511. J Gen Physiol. 1999. PMID: 10498670 Free PMC article.
-
Adaptive plasticity during the development of colour vision.Prog Retin Eye Res. 2005 Jul;24(4):521-36. doi: 10.1016/j.preteyeres.2005.01.002. Prog Retin Eye Res. 2005. PMID: 15845347 Review.
-
The involvement of glutamate-gated channels in negative feedback from horizontal cells to cones.Prog Brain Res. 2005;147:219-29. doi: 10.1016/S0079-6123(04)47017-4. Prog Brain Res. 2005. PMID: 15581709 Review.
Cited by
-
High Contrast Allows the Retina to Compute More Than Just Contrast.Front Cell Neurosci. 2021 Jan 15;14:595193. doi: 10.3389/fncel.2020.595193. eCollection 2020. Front Cell Neurosci. 2021. PMID: 33519381 Free PMC article.
-
Simple model for encoding natural images by retinal ganglion cells with nonlinear spatial integration.PLoS Comput Biol. 2022 Mar 8;18(3):e1009925. doi: 10.1371/journal.pcbi.1009925. eCollection 2022 Mar. PLoS Comput Biol. 2022. PMID: 35259159 Free PMC article.
-
Heteromeric KV2/KV8.2 Channels Mediate Delayed Rectifier Potassium Currents in Primate Photoreceptors.J Neurosci. 2018 Apr 4;38(14):3414-3427. doi: 10.1523/JNEUROSCI.2440-17.2018. Epub 2018 Feb 26. J Neurosci. 2018. PMID: 29483285 Free PMC article.
-
Regional tuning of photoreceptor adaptation in the primate retina.Nat Commun. 2024 Oct 12;15(1):8821. doi: 10.1038/s41467-024-53061-3. Nat Commun. 2024. PMID: 39394185 Free PMC article.
-
The Roles of Rods, Cones, and Melanopsin in Photoresponses of M4 Intrinsically Photosensitive Retinal Ganglion Cells (ipRGCs) and Optokinetic Visual Behavior.Front Cell Neurosci. 2018 Jul 12;12:203. doi: 10.3389/fncel.2018.00203. eCollection 2018. Front Cell Neurosci. 2018. PMID: 30050414 Free PMC article.
References
-
- Fain GL, Matthews HR, Cornwall MC, Koutalos Y. Adaptation in vertebrate photoreceptors. Physiol Rev. 2001;81(1):117–51. - PubMed
-
- van der Schaaf A, van Hateren JH. Modelling the power spectra of natural images: statistics and information. Vision Res. 1996;36(17):2759–70. - PubMed
-
- Balboa RM, Grzywacz NM. Power spectra and distribution of contrasts of natural images from different habitats. Vision Res. 2003;43(24):2527–37. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
