

Transcriptional repression of frequency by the IEC-1-INO80 complex is required for normal Neurospora circadian clock function
- PMID: 28403234
- PMCID: PMC5406019
- DOI: 10.1371/journal.pgen.1006732
Transcriptional repression of frequency by the IEC-1-INO80 complex is required for normal Neurospora circadian clock function
Abstract
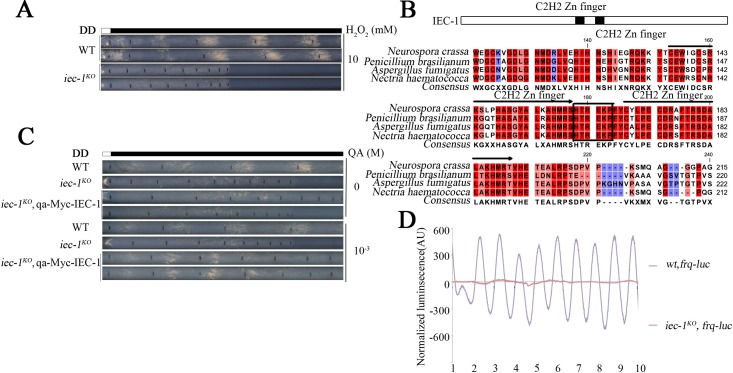
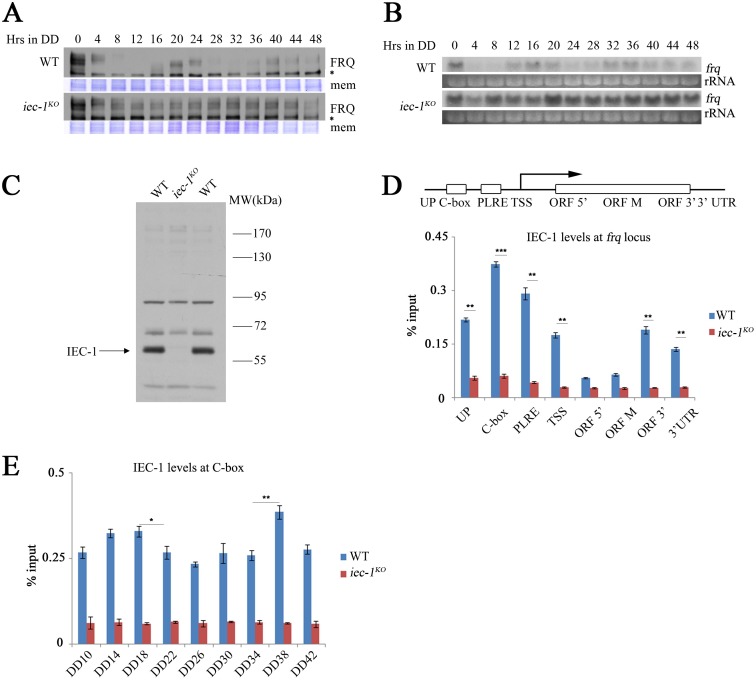
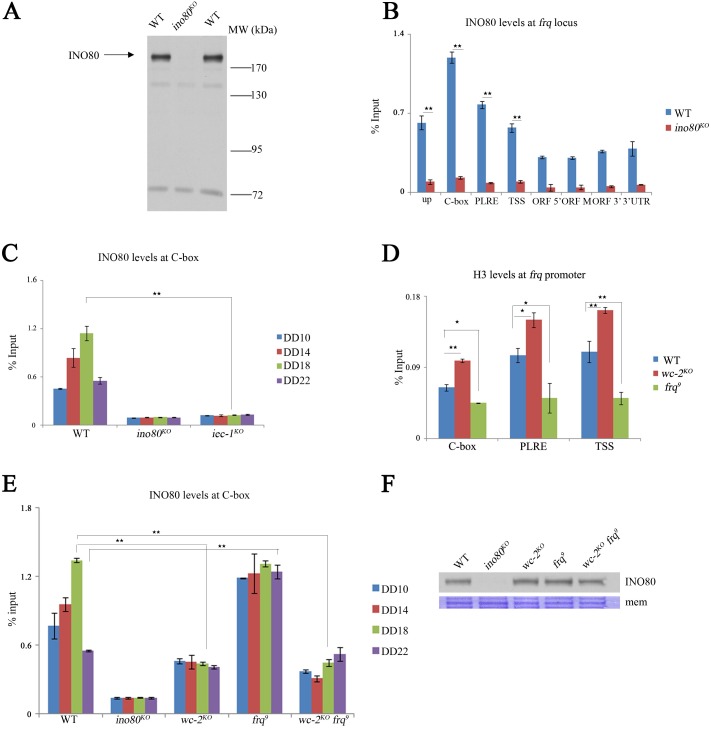
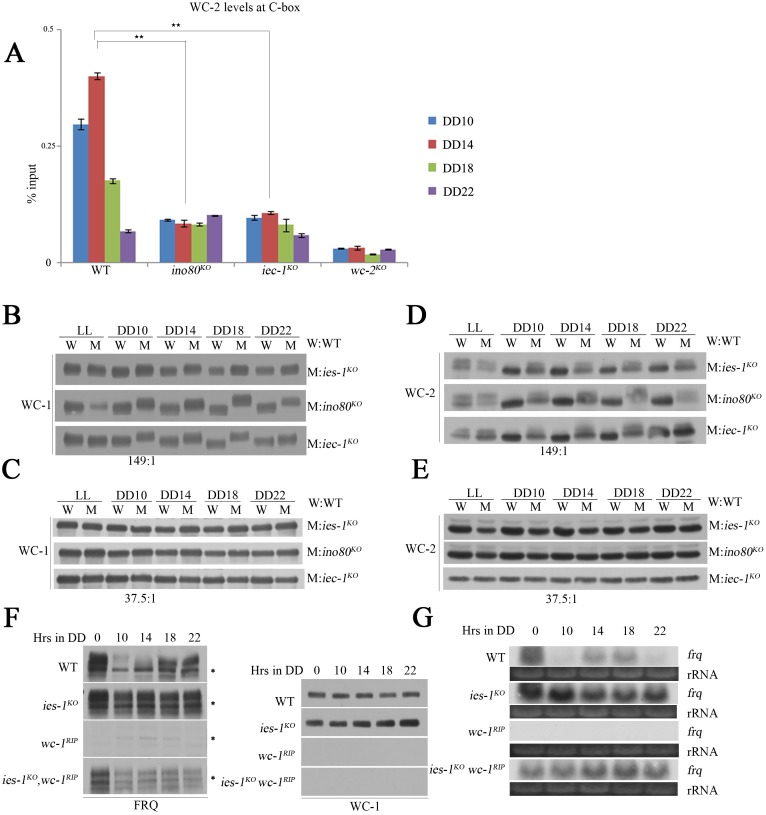
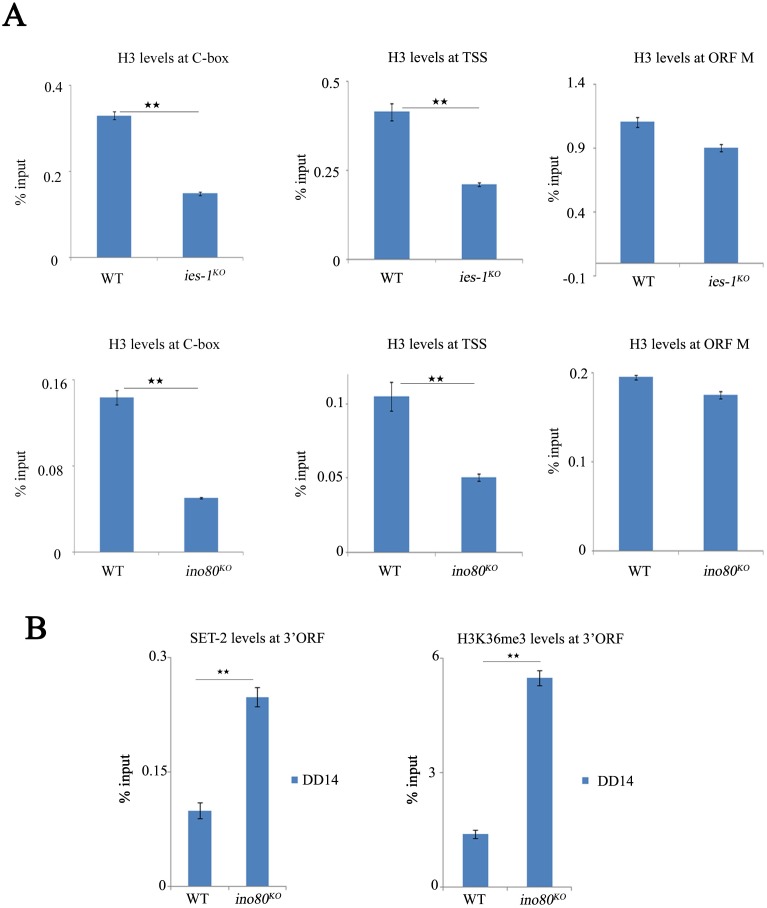
Rhythmic activation and repression of the frequency (frq) gene are essential for normal function of the Neurospora circadian clock. WHITE COLLAR (WC) complex, the positive element of the Neurospora circadian system, is responsible for stimulation of frq transcription. We report that a C2H2 finger domain-containing protein IEC-1 and its associated chromatin remodeling complex INO80 play important roles in normal Neurospora circadian clock function. In iec-1KO strains, circadian rhythms are abolished, and the frq transcript levels are increased compared to that of the wild-type strain. Similar results are observed in mutant strains of the INO80 subunits. Furthermore, ChIP data show that recruitment of the INO80 complex to the frq promoter is IEC-1-dependent. WC-mediated transcription of frq contributes to the rhythmic binding of the INO80 complex at the frq promoter. As demonstrated by ChIP analysis, the INO80 complex is required for the re-establishment of the dense chromatin environment at the frq promoter. In addition, WC-independent frq transcription is present in ino80 mutants. Altogether, our data indicate that the INO80 complex suppresses frq transcription by re-assembling the suppressive mechanisms at the frq promoter after transcription of frq.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Transcription factor CBF-1 is critical for circadian gene expression by modulating WHITE COLLAR complex recruitment to the frq locus.PLoS Genet. 2018 Sep 12;14(9):e1007570. doi: 10.1371/journal.pgen.1007570. eCollection 2018 Sep. PLoS Genet. 2018. PMID: 30208021 Free PMC article.
-
Suppression of WHITE COLLAR-independent frequency Transcription by Histone H3 Lysine 36 Methyltransferase SET-2 Is Necessary for Clock Function in Neurospora.J Biol Chem. 2016 May 20;291(21):11055-63. doi: 10.1074/jbc.M115.711333. Epub 2016 Mar 21. J Biol Chem. 2016. PMID: 27002152 Free PMC article.
-
Role for Protein Kinase A in the Neurospora Circadian Clock by Regulating White Collar-Independent frequency Transcription through Phosphorylation of RCM-1.Mol Cell Biol. 2015 Jun;35(12):2088-102. doi: 10.1128/MCB.00709-14. Epub 2015 Apr 6. Mol Cell Biol. 2015. PMID: 25848091 Free PMC article.
-
The molecular workings of the Neurospora biological clock.Novartis Found Symp. 2003;253:184-98; discussion 102-9, 198-202, 281-4. Novartis Found Symp. 2003. PMID: 14712922 Review.
-
The neurospora circadian system.J Biol Rhythms. 2004 Oct;19(5):414-24. doi: 10.1177/0748730404269116. J Biol Rhythms. 2004. PMID: 15534321 Review.
Cited by
-
NC2 complex is a key factor for the activation of catalase-3 transcription by regulating H2A.Z deposition.Nucleic Acids Res. 2020 Sep 4;48(15):8332-8348. doi: 10.1093/nar/gkaa552. Nucleic Acids Res. 2020. PMID: 32633757 Free PMC article.
-
The Frq-Frh Complex Light-Dependently Delays Sfl1-Induced Microsclerotia Formation in Verticillium dahliae.J Fungi (Basel). 2023 Jul 4;9(7):725. doi: 10.3390/jof9070725. J Fungi (Basel). 2023. PMID: 37504714 Free PMC article.
-
A global search for novel transcription factors impacting the Neurospora crassa circadian clock.G3 (Bethesda). 2021 Jun 17;11(6):jkab100. doi: 10.1093/g3journal/jkab100. G3 (Bethesda). 2021. PMID: 33792687 Free PMC article.
-
Transcription factor VIB-1 activates catalase-3 expression by promoting PIC assembly in Neurospora crassa.Nucleic Acids Res. 2025 Feb 27;53(5):gkaf174. doi: 10.1093/nar/gkaf174. Nucleic Acids Res. 2025. PMID: 40087884 Free PMC article.
-
Transcriptional rewiring of an evolutionarily conserved circadian clock.EMBO J. 2024 May;43(10):2015-2034. doi: 10.1038/s44318-024-00088-3. Epub 2024 Apr 16. EMBO J. 2024. PMID: 38627599 Free PMC article.
References
-
- Dunlap JC (1999) Molecular Bases for Circadian Clocks. Cell 96: 271–290. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
