

Mechanisms Mediating Functional Hyperemia in the Brain
- PMID: 28403673
- PMCID: PMC5757525
- DOI: 10.1177/1073858417703033
Mechanisms Mediating Functional Hyperemia in the Brain
Abstract
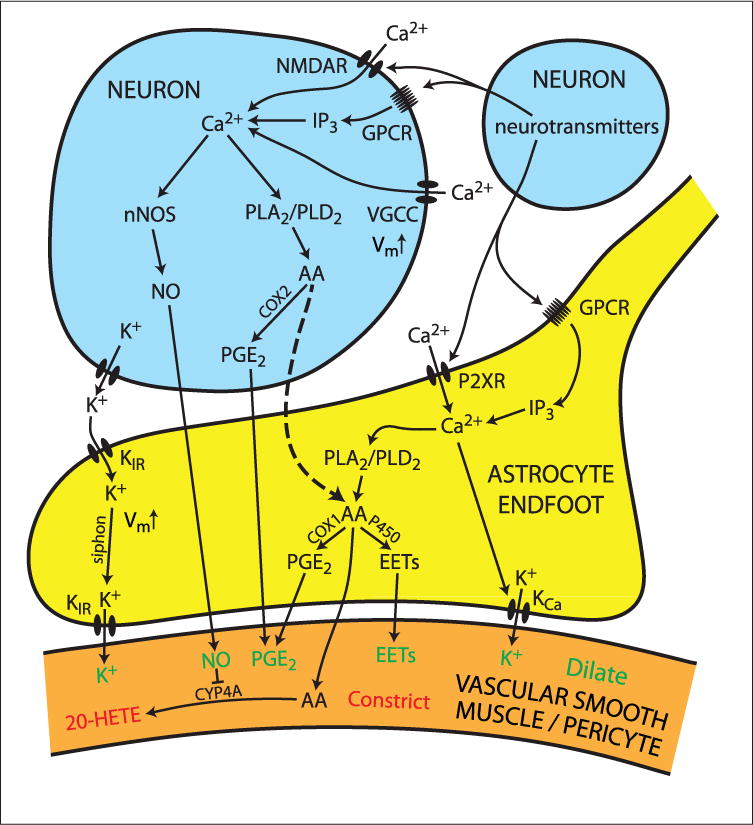
Neuronal activity within the brain evokes local increases in blood flow, a response termed functional hyperemia. This response ensures that active neurons receive sufficient oxygen and nutrients to maintain tissue function and health. In this review, we discuss the functions of functional hyperemia, the types of vessels that generate the response, and the signaling mechanisms that mediate neurovascular coupling, the communication between neurons and blood vessels. Neurovascular coupling signaling is mediated primarily by the vasoactive metabolites of arachidonic acid (AA), by nitric oxide, and by K+. While much is known about these pathways, many contentious issues remain. We highlight two controversies, the role of glial cell Ca2+ signaling in mediating neurovascular coupling and the importance of capillaries in generating functional hyperemia. We propose signaling pathways that resolve these controversies. In this scheme, capillary dilations are generated by Ca2+ increases in astrocyte endfeet, leading to production of AA metabolites. In contrast, arteriole dilations are generated by Ca2+ increases in neurons, resulting in production of nitric oxide and AA metabolites. Arachidonic acid from neurons also diffuses into astrocyte endfeet where it is converted into additional vasoactive metabolites. While this scheme resolves several discrepancies in the field, many unresolved challenges remain and are discussed in the final section of the review.
Keywords: EETs; PGE2; arachidonic acid; astrocyte; cerebral blood flow; functional hyperemia; neurovascular coupling.
Conflict of interest statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Figures


References
-
- Akgoren N, Dalgaard P, Lauritzen M. Cerebral blood flow increases evoked by electrical stimulation of rat cerebellar cortex: relation to excitatory synaptic activity and nitric oxide synthesis. Brain Res. 1996;710:204–14. - PubMed
-
- Alkayed NJ, Birks EK, Hudetz AG, Roman RJ, Henderson L, Harder DR. Inhibition of brain P-450 arachidonic acid epoxygenase decreases baseline cerebral blood flow. Am J Physiol. 1996a;271:H1541–6. - PubMed
-
- Alkayed NJ, Narayanan J, Gebremedhin D, Medhora M, Roman RJ, Harder DR. Molecular characterization of an arachidonic acid epoxygenase in rat brain astrocytes. Stroke. 1996b;27:971–9. - PubMed
-
- Anderson AW, Marois R, Colson ER, Peterson BS, Duncan CC, Ehrenkranz RA, et al. Neonatal auditory activation detected by functional magnetic resonance imaging. Magn Reson Imaging. 2001;19:1–5. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
