

Surfactant inhibits ATP-induced release of interleukin-1β via nicotinic acetylcholine receptors
- PMID: 28404637
- PMCID: PMC5454502
- DOI: 10.1194/jlr.M071506
Surfactant inhibits ATP-induced release of interleukin-1β via nicotinic acetylcholine receptors
Abstract
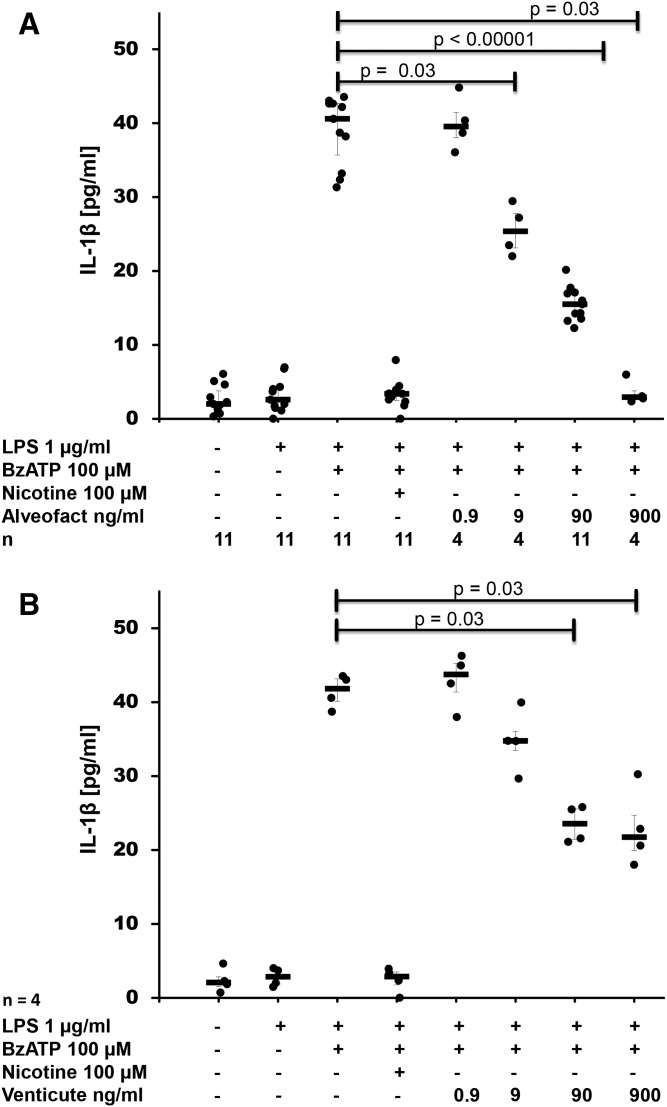
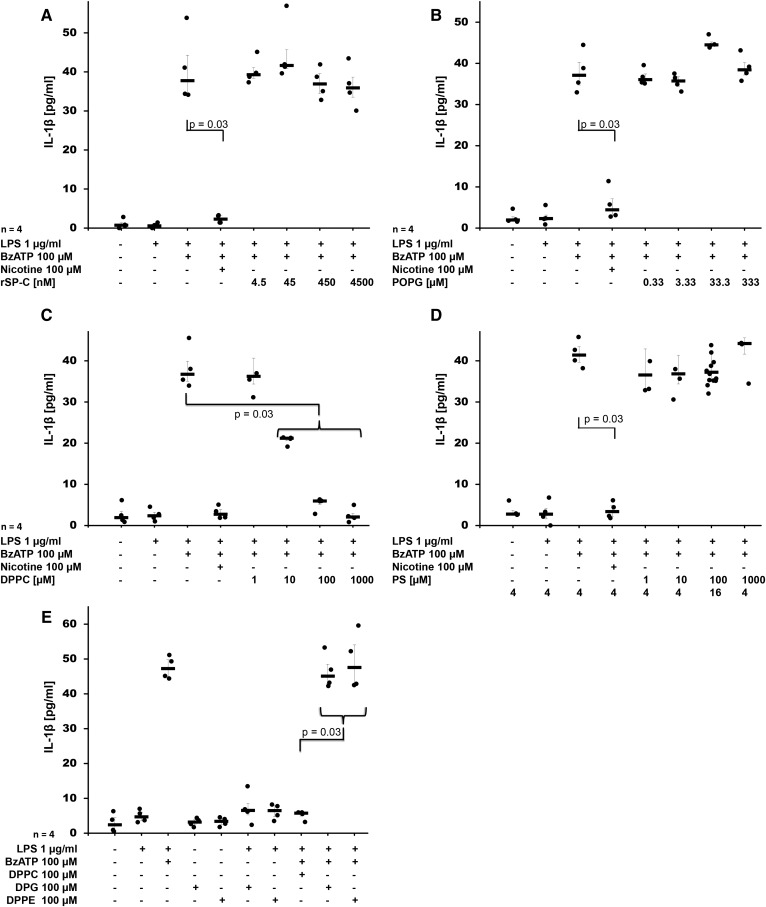
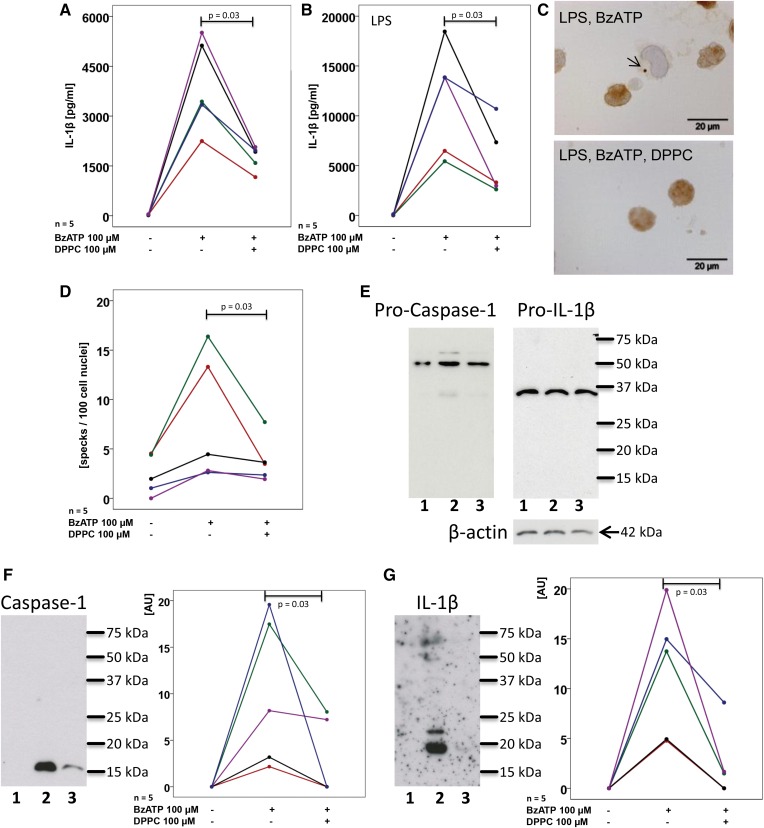
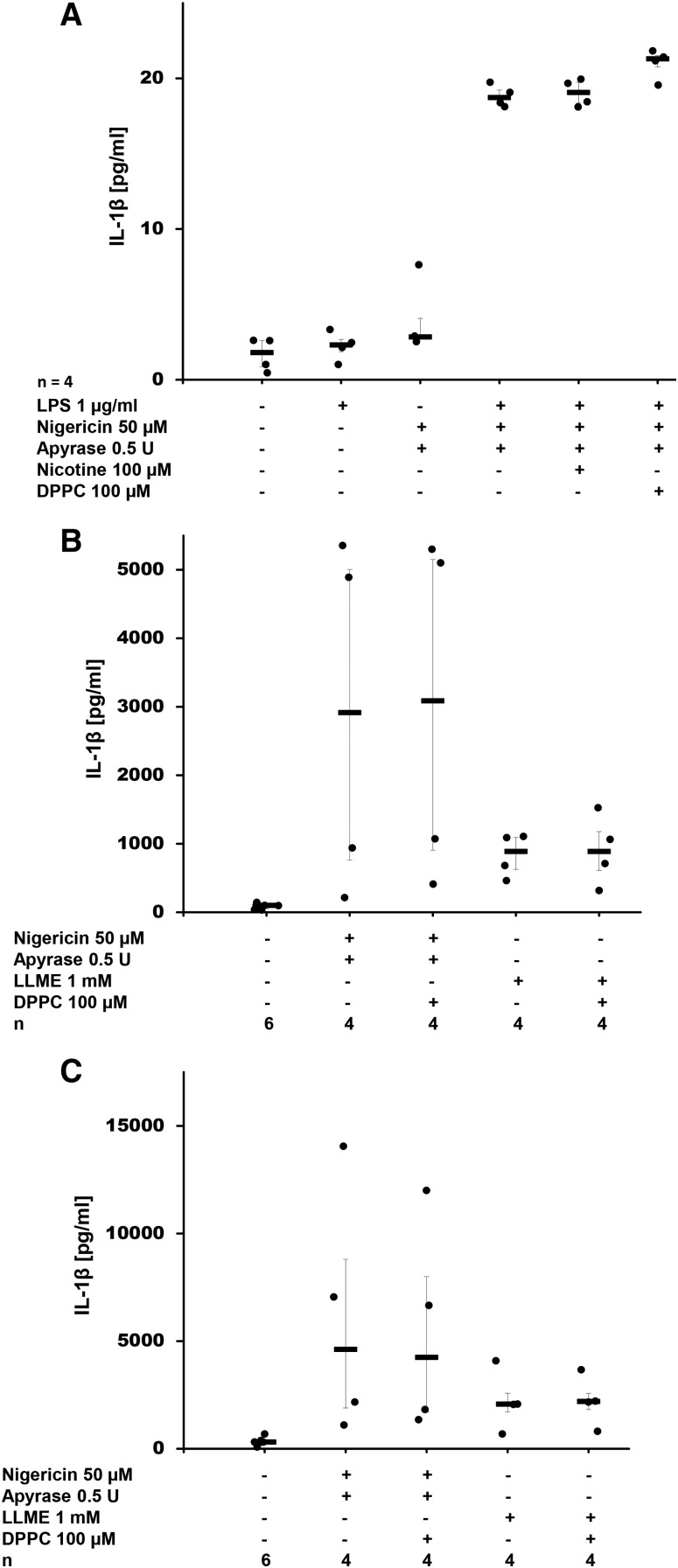
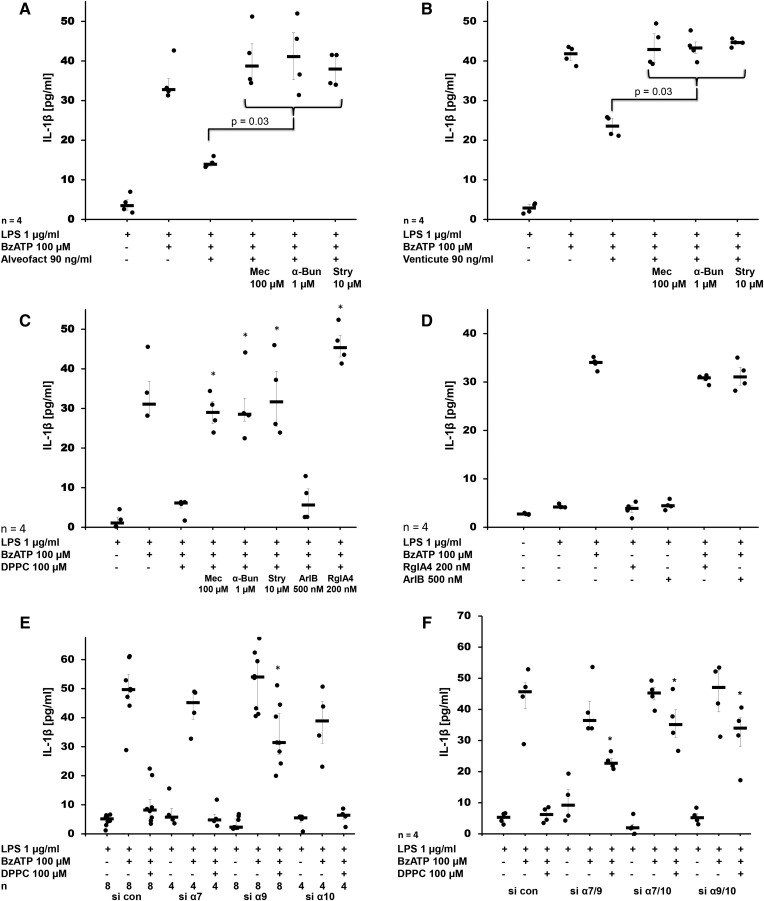
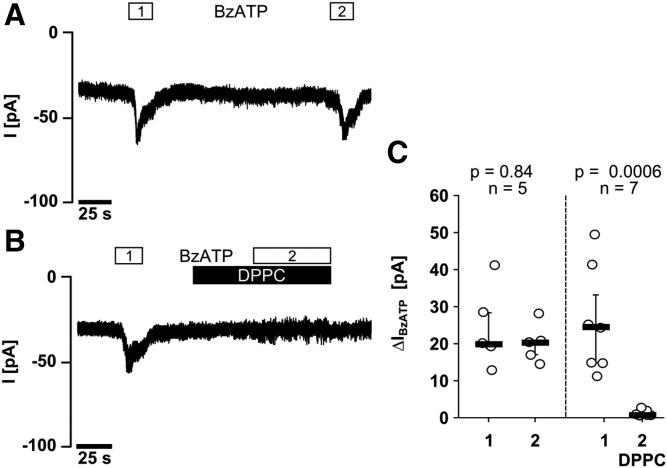
Interleukin (IL)-1β is a potent pro-inflammatory cytokine of innate immunity involved in host defense. High systemic IL-1β levels, however, cause life-threatening inflammatory diseases, including systemic inflammatory response syndrome. In response to various danger signals, the pro-form of IL-1β is synthesized and stays in the cytoplasm unless a second signal, such as extracellular ATP, activates the inflammasome, which enables processing and release of mature IL-1β. As pulmonary surfactant is known for its anti-inflammatory properties, we hypothesize that surfactant inhibits ATP-induced release of IL-1β. Lipopolysaccharide-primed monocytic U937 cells were stimulated with an ATP analog in the presence of natural or synthetic surfactant composed of recombinant surfactant protein (rSP)-C, palmitoylphosphatidylglycerol, and dipalmitoylphosphatidylcholine (DPPC). Both surfactant preparations dose-dependently inhibited IL-1β release from U937 cells. DPPC was the active constituent of surfactant, whereas rSP-C and palmitoylphosphatidylglycerol were inactive. DPPC was also effective in primary mononuclear leukocytes isolated from human blood. Experiments with nicotinic antagonists, siRNA technology, and patch-clamp experiments suggested that stimulation of nicotinic acetylcholine receptors (nAChRs) containing subunit α9 results in a complete inhibition of the ion channel function of ATP receptor, P2X7. In conclusion, the surfactant constituent, DPPC, efficiently inhibits ATP-induced inflammasome activation and maturation of IL-1β in human monocytes by a mechanism involving nAChRs.
Keywords: CHRNA10; CHRNA7; CHRNA9; dipalmitoylphosphatidylcholine; inflammasome; monocyte; phosphatidylcholine; purinergic receptor P2X 7.
Copyright © 2017 by the American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Hiemstra P. S. 2007. The role of epithelial beta-defensins and cathelicidins in host defense of the lung. Exp. Lung Res. 33: 537–542. - PubMed
-
- Alkhouri H., Poppinga W. J., Tania N. P., Ammit A., and Schuliga M.. 2014. Regulation of pulmonary inflammation by mesenchymal cells. Pulm. Pharmacol. Ther. 29: 156–165. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
