

Preprotein mature domains contain translocase targeting signals that are essential for secretion
- PMID: 28404644
- PMCID: PMC5412566
- DOI: 10.1083/jcb.201609022
Preprotein mature domains contain translocase targeting signals that are essential for secretion
Abstract
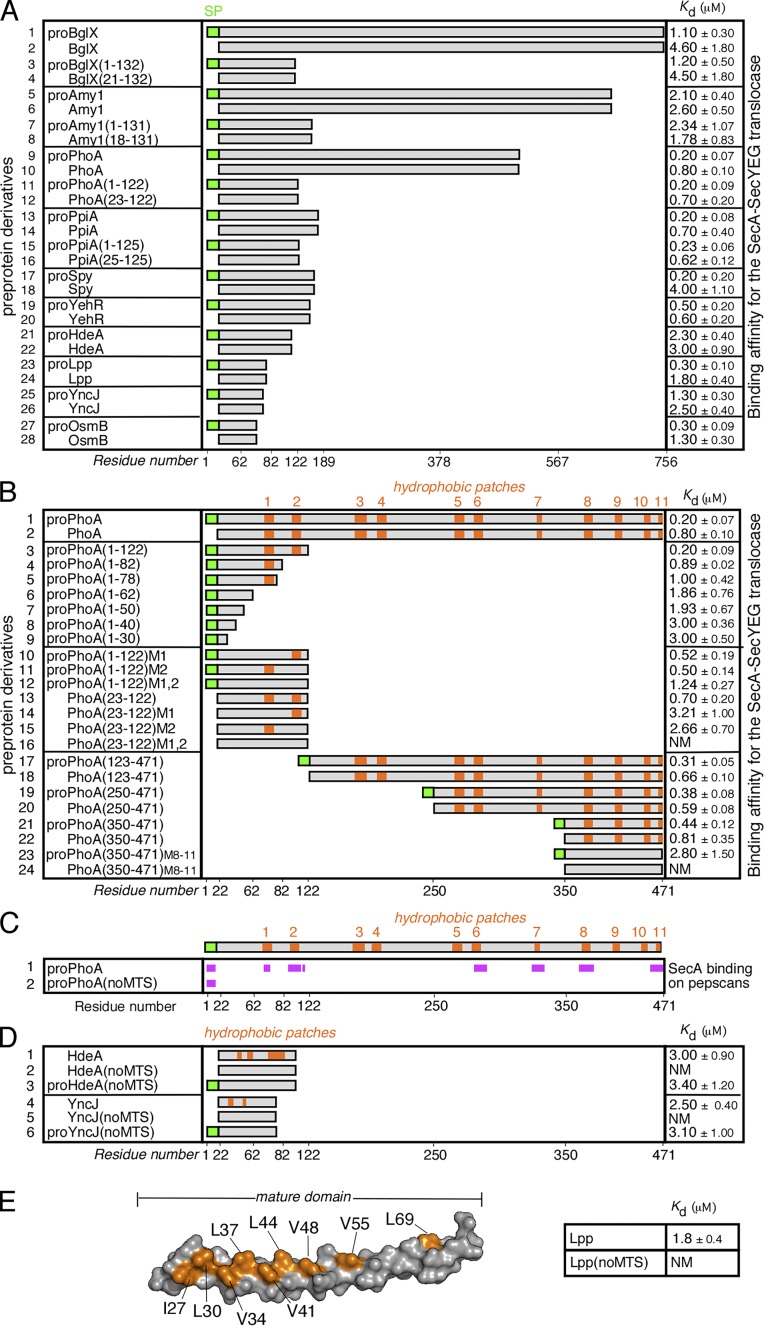
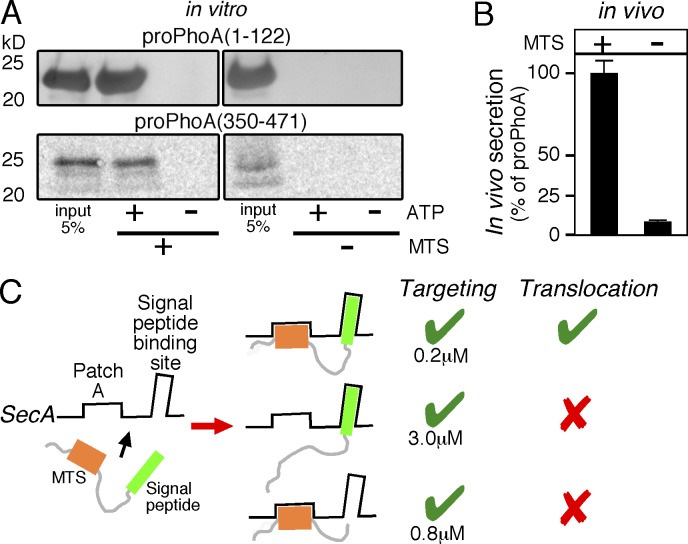
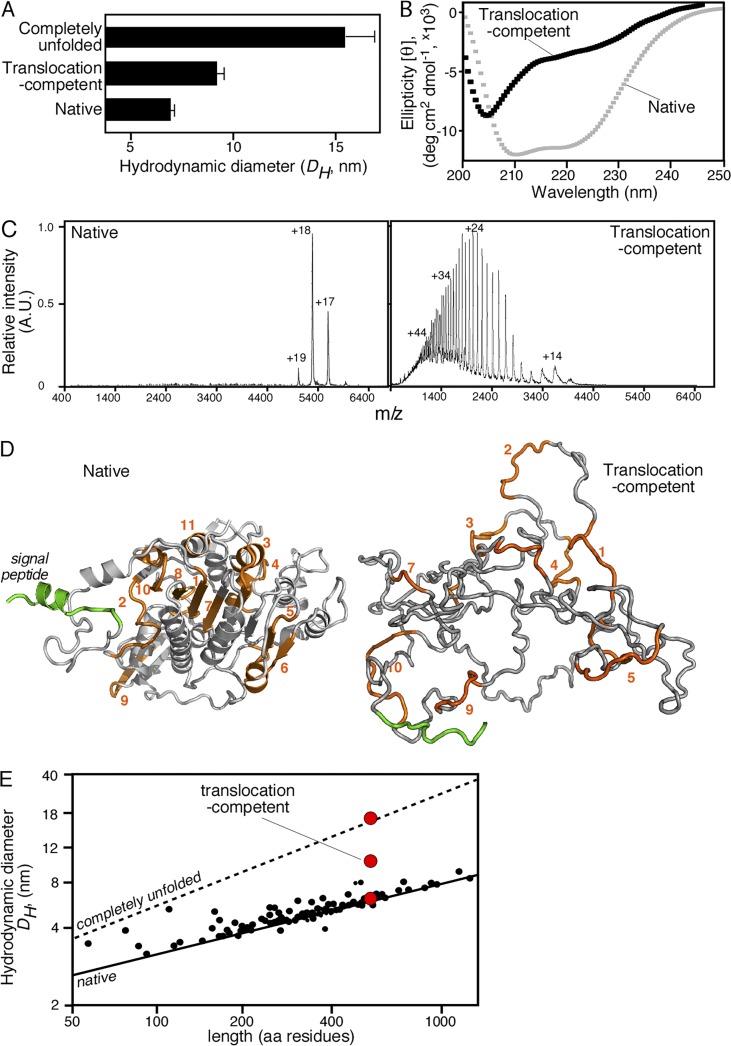
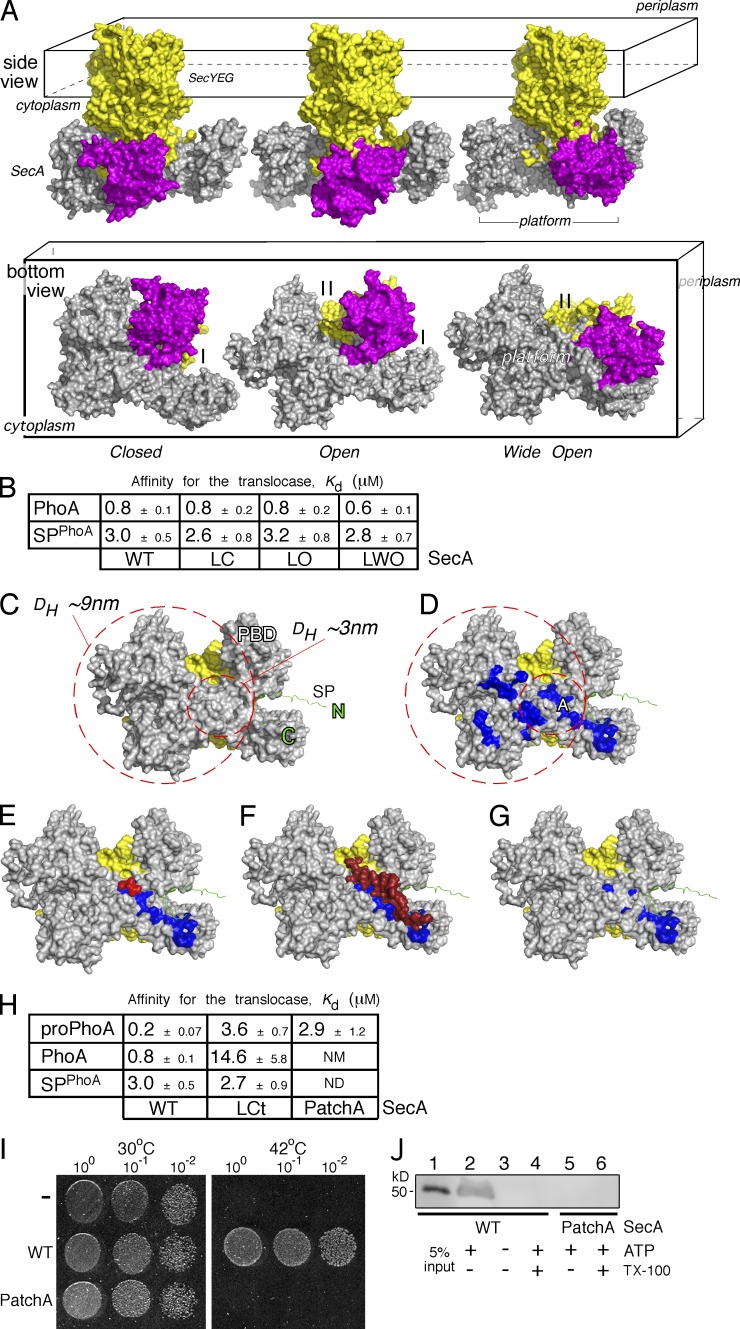
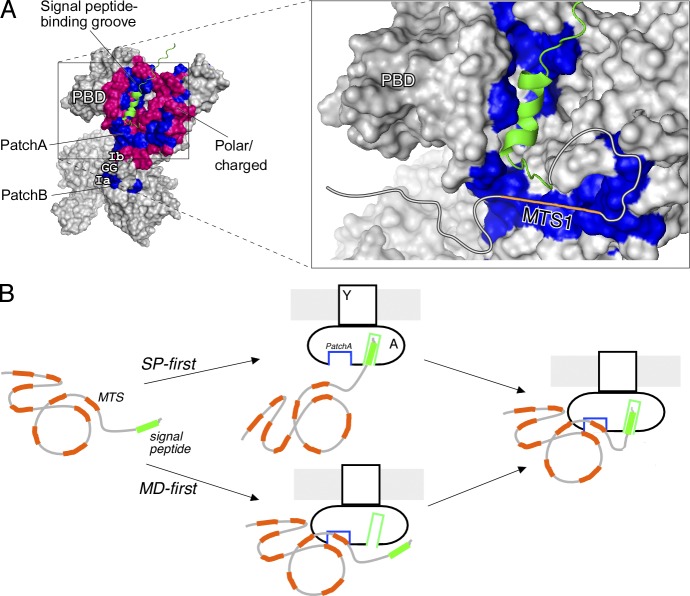
Secretory proteins are only temporary cytoplasmic residents. They are typically synthesized as preproteins, carrying signal peptides N-terminally fused to their mature domains. In bacteria secretion largely occurs posttranslationally through the membrane-embedded SecA-SecYEG translocase. Upon crossing the plasma membrane, signal peptides are cleaved off and mature domains reach their destinations and fold. Targeting to the translocase is mediated by signal peptides. The role of mature domains in targeting and secretion is unclear. We now reveal that mature domains harbor their own independent targeting signals (mature domain targeting signals [MTSs]). These are multiple, degenerate, interchangeable, linear or 3D hydrophobic stretches that become available because of the unstructured states of targeting-competent preproteins. Their receptor site on the cytoplasmic face of the SecYEG-bound SecA is also of hydrophobic nature and is located adjacent to the signal peptide cleft. Both the preprotein MTSs and their receptor site on SecA are essential for protein secretion. Evidently, mature domains have their own previously unsuspected distinct roles in preprotein targeting and secretion.
© 2017 Chatzi et al.
Figures
References
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
