

Ecology of cold environments: new insights of bacterial metabolic adaptation through an integrated genomic-phenomic approach
- PMID: 28404986
- PMCID: PMC5429795
- DOI: 10.1038/s41598-017-00876-4
Ecology of cold environments: new insights of bacterial metabolic adaptation through an integrated genomic-phenomic approach
Abstract
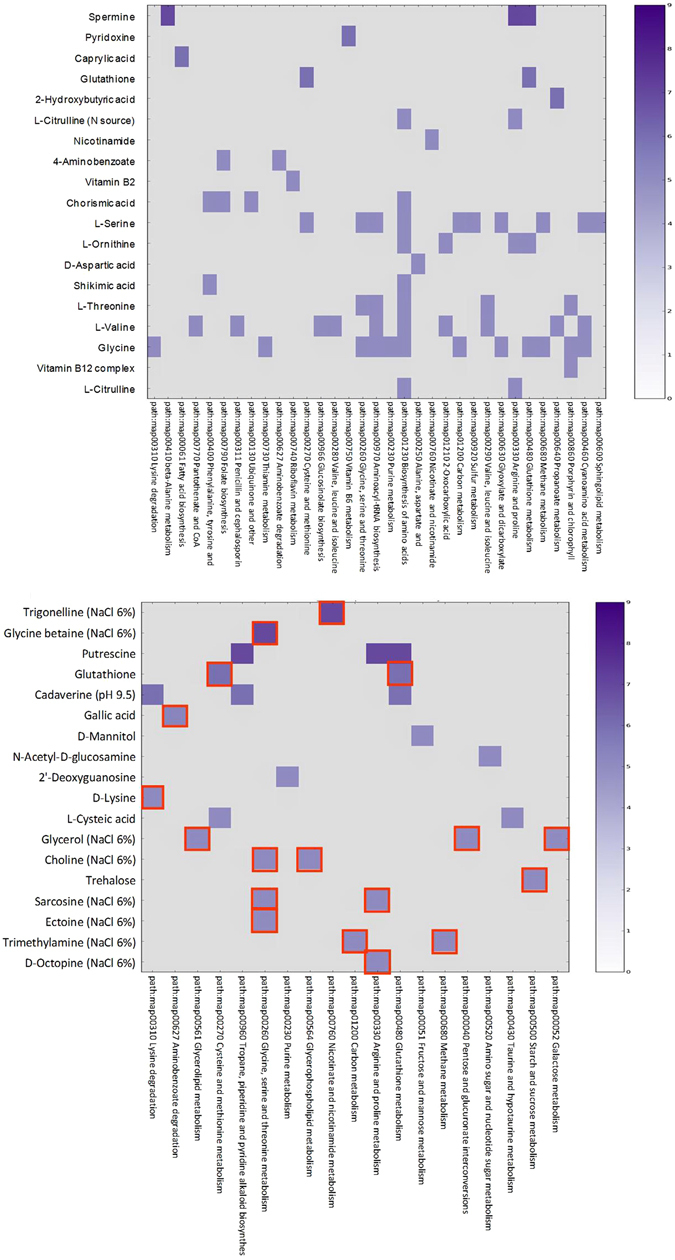
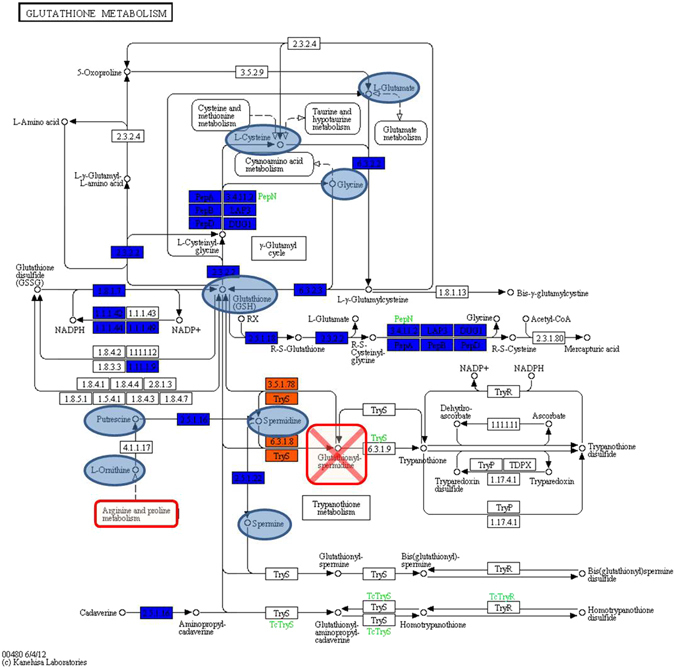
Cold environments dominate Earth's biosphere, hosting complex microbial communities with the ability to thrive at low temperatures. However, the underlying molecular mechanisms and the metabolic pathways involved in bacterial cold-adaptation mechanisms are still not fully understood. Herein, we assessed the metabolic features of the Antarctic bacterium Pseudoalteromonas haloplanktis TAC125 (PhTAC125), a model organism for cold-adaptation, at both 4 °C and 15 °C, by integrating genomic and phenomic (high-throughput phenotyping) data and comparing the obtained results to the taxonomically related Antarctic bacterium Pseudoalteromonas sp. TB41 (PspTB41). Although the genome size of PspTB41 is considerably larger than PhTAC125, the higher number of genes did not reflect any higher metabolic versatility at 4 °C as compared to PhTAC125. Remarkably, protein S-thiolation regulated by glutathione and glutathionylspermidine appeared to be a new possible mechanism for cold adaptation in PhTAC125. More in general, this study represents an example of how 'multi-omic' information might potentially contribute in filling the gap between genotypic and phenotypic features related to cold-adaptation mechanisms in bacteria.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
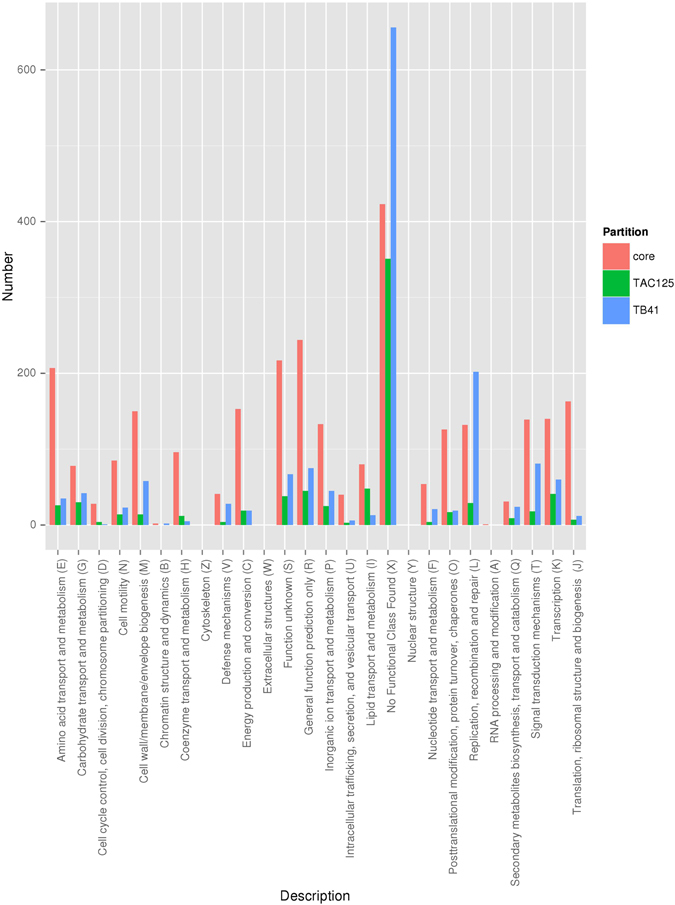
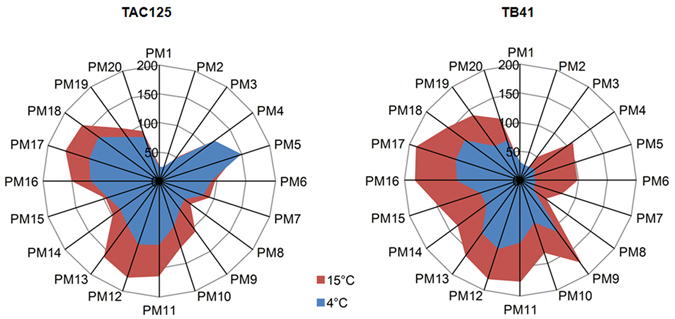
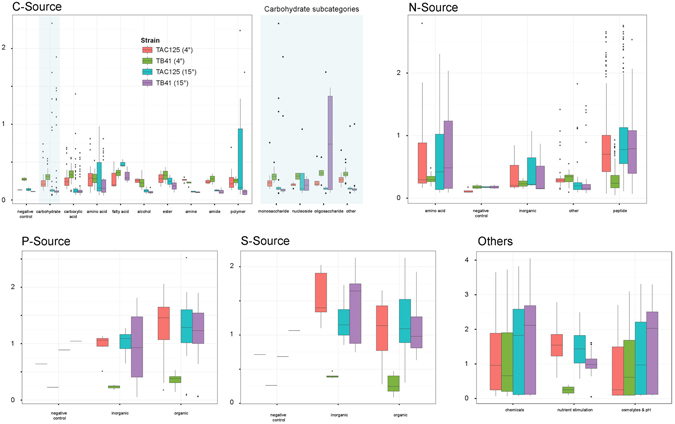
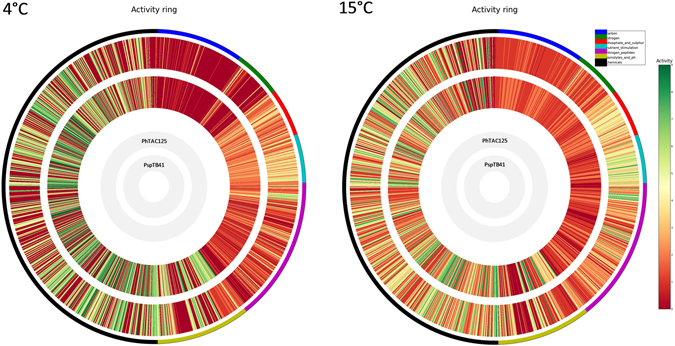
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
