

Glucose uptake in Azotobacter vinelandii occurs through a GluP transporter that is under the control of the CbrA/CbrB and Hfq-Crc systems
- PMID: 28404995
- PMCID: PMC5429807
- DOI: 10.1038/s41598-017-00980-5
Glucose uptake in Azotobacter vinelandii occurs through a GluP transporter that is under the control of the CbrA/CbrB and Hfq-Crc systems
Abstract
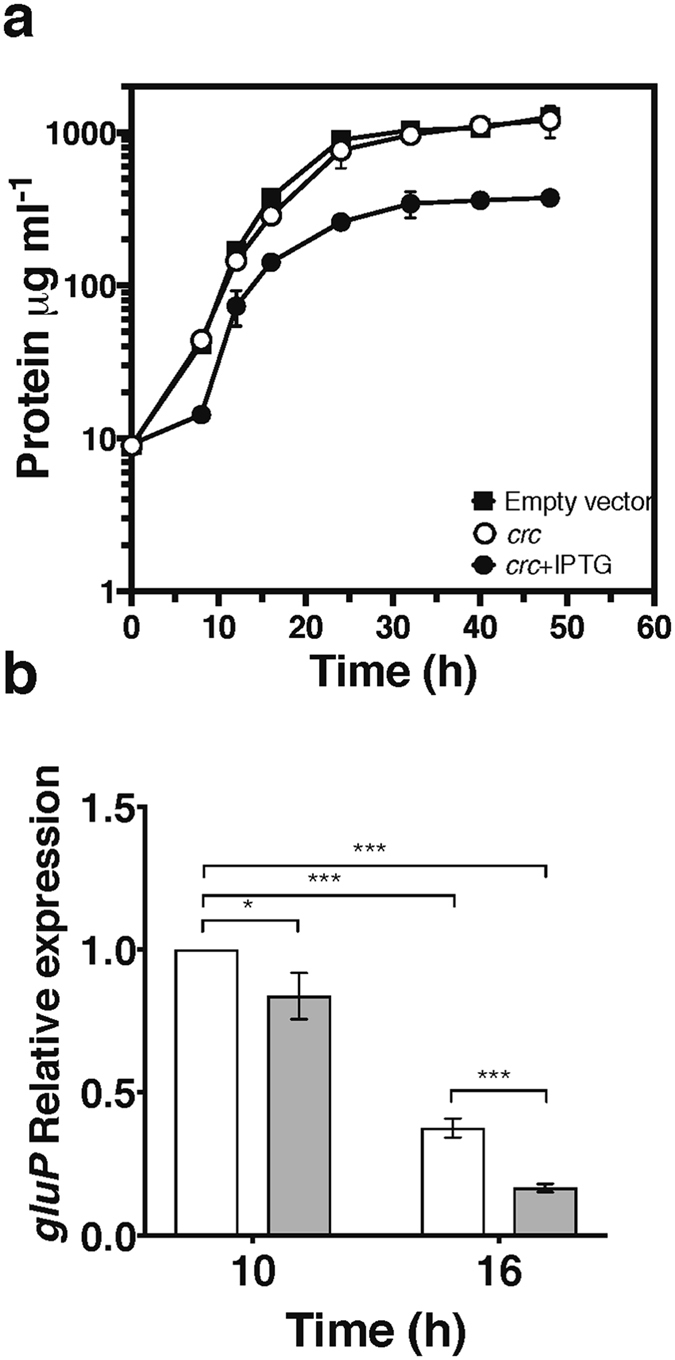
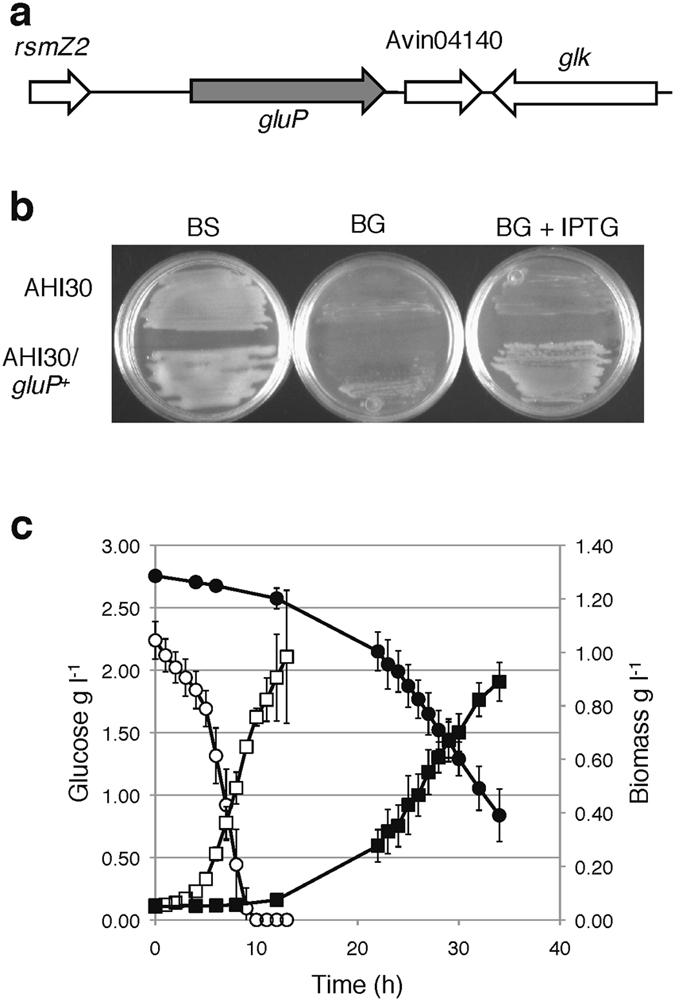
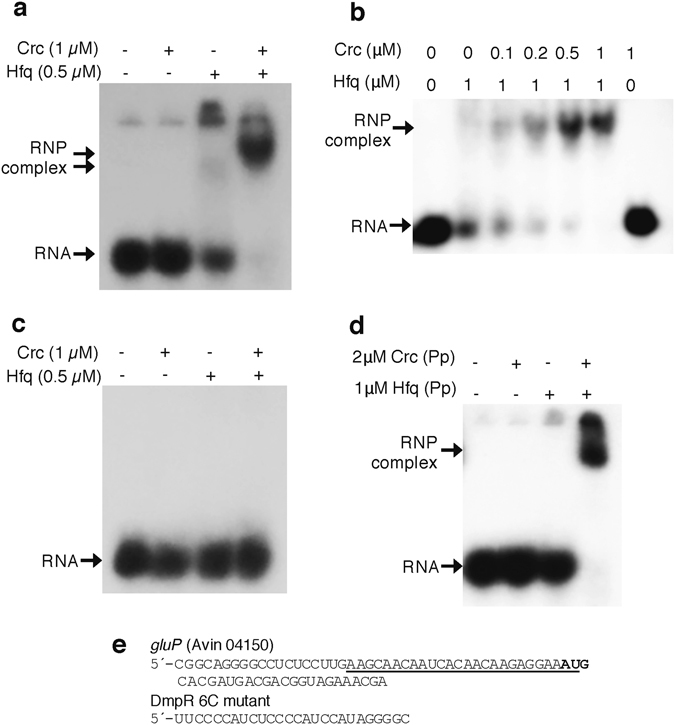
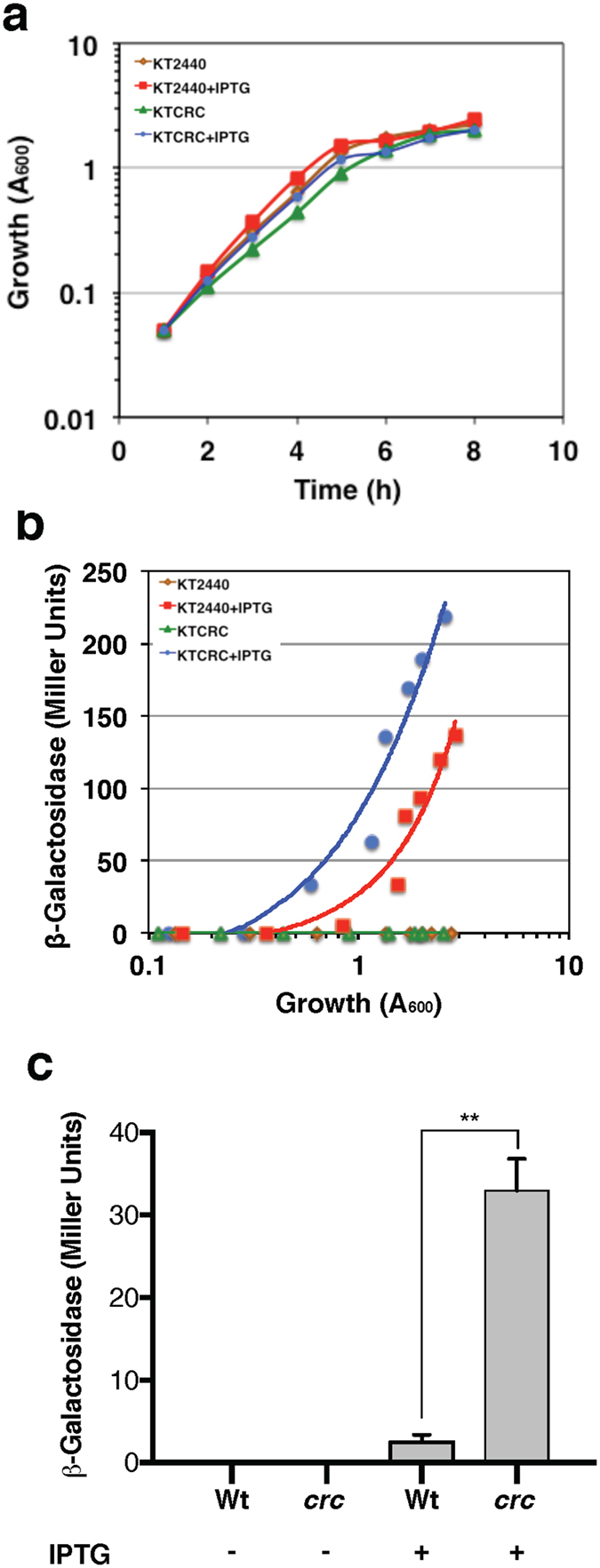
Azotobacter vinelandii, a strict aerobic, nitrogen fixing bacterium in the Pseudomonadaceae family, exhibits a preferential use of acetate over glucose as a carbon source. In this study, we show that GluP (Avin04150), annotated as an H+-coupled glucose-galactose symporter, is the glucose transporter in A. vinelandii. This protein, which is widely distributed in bacteria and archaea, is uncommon in Pseudomonas species. We found that expression of gluP was under catabolite repression control thorugh the CbrA/CbrB and Crc/Hfq regulatory systems, which were functionally conserved between A. vinelandii and Pseudomonas species. While the histidine kinase CbrA was essential for glucose utilization, over-expression of the Crc protein arrested cell growth when glucose was the sole carbon source. Crc and Hfq proteins from either A. vinelandii or P. putida could form a stable complex with an RNA A-rich Hfq-binding motif present in the leader region of gluP mRNA. Moreover, in P. putida, the gluP A-rich Hfq-binding motif was functional and promoted translational inhibition of a lacZ reporter gene. The fact that gluP is not widely distributed in the Pseudomonas genus but is under control of the CbrA/CbrB and Crc/Hfq systems demonstrates the relevance of these systems in regulating metabolism in the Pseudomonadaceae family.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
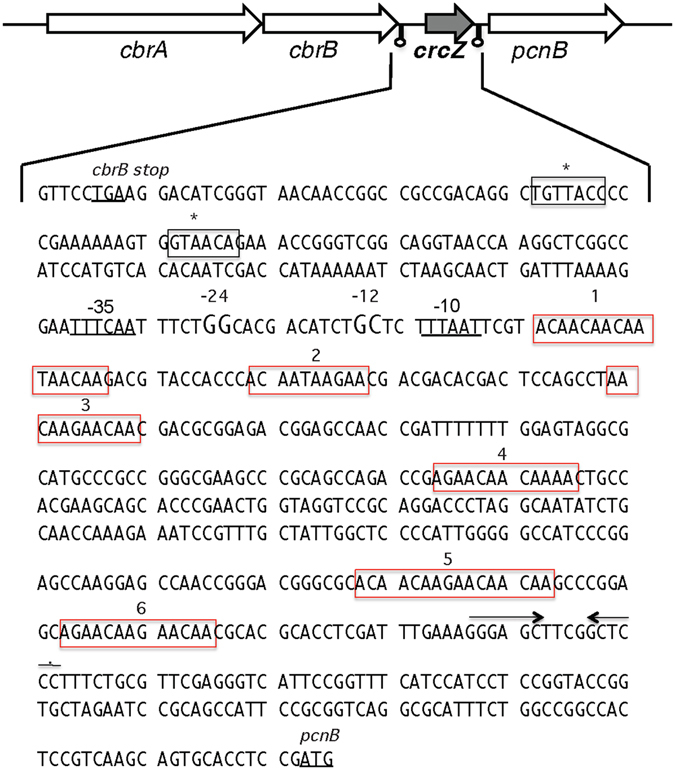
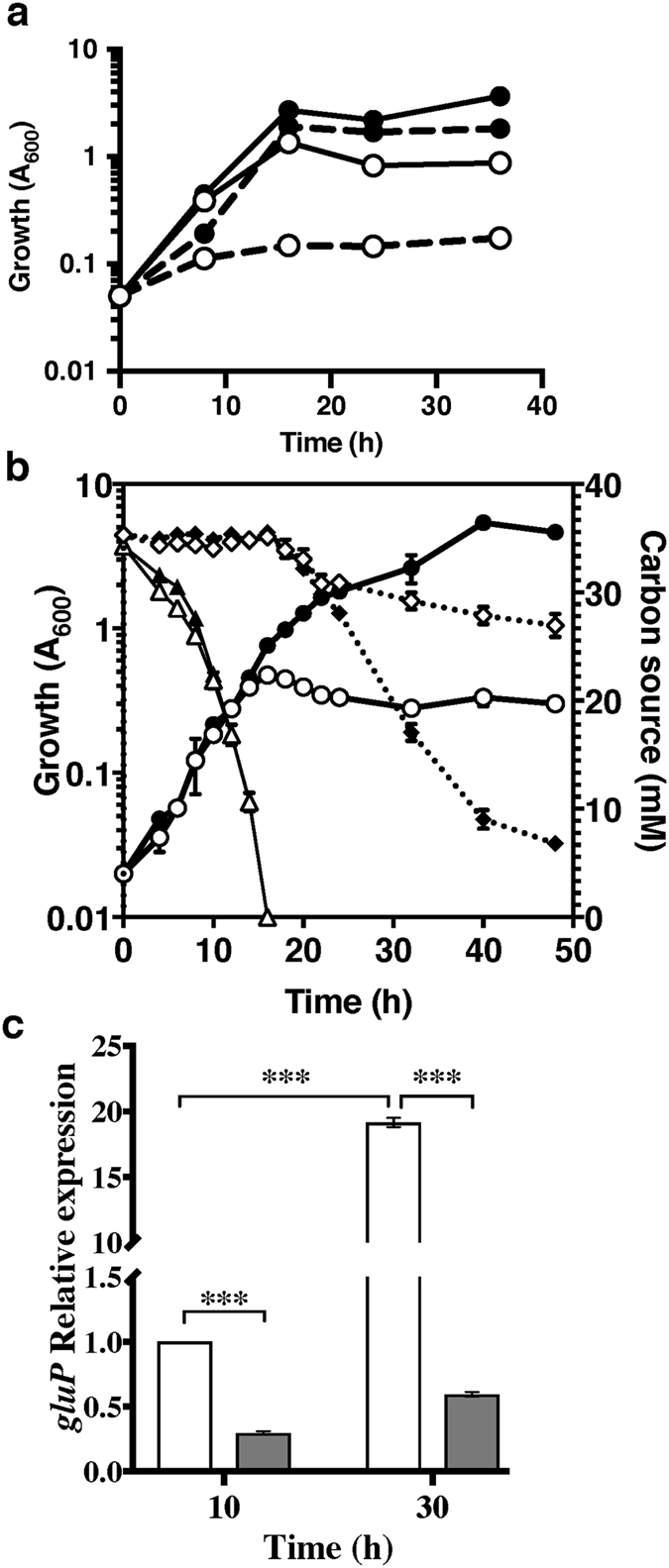
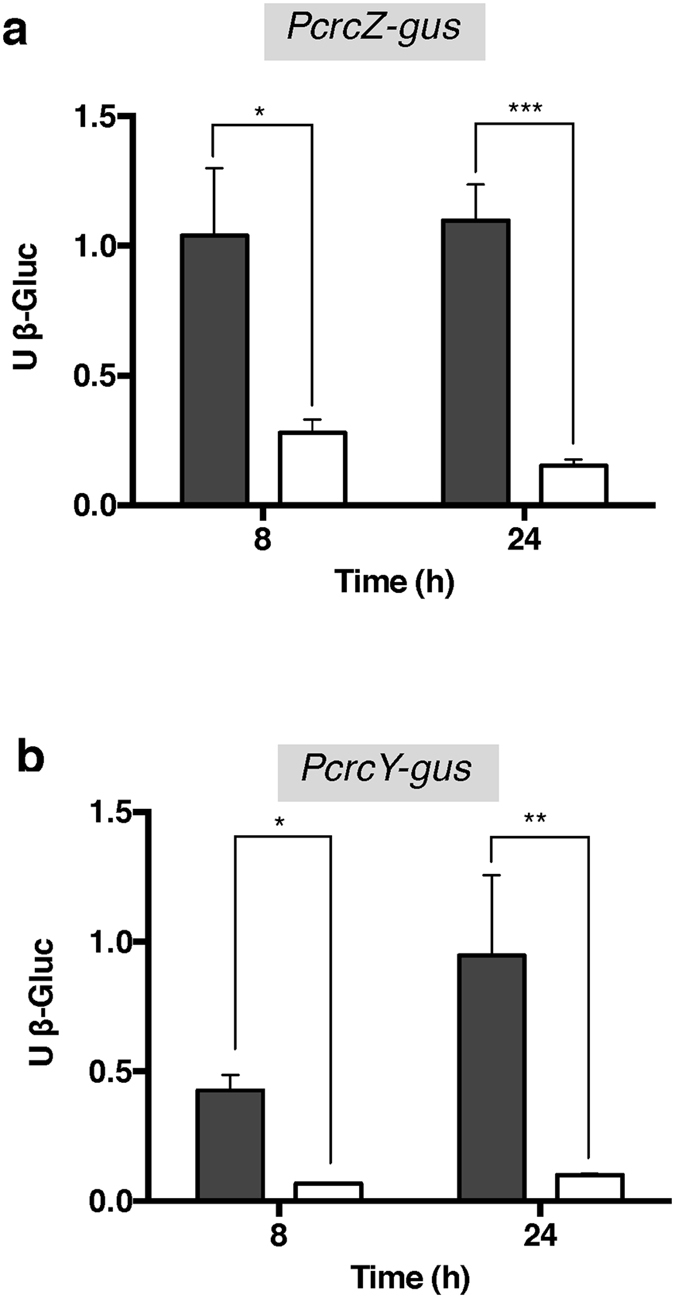
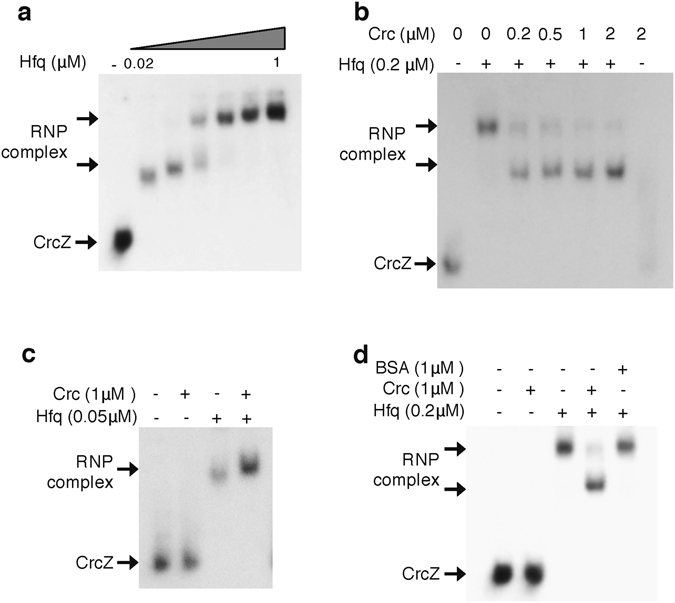
References
-
- Kennedy, C., Rudnick, P., MacDonald, T. & Melton, T. Genus Azotobacter.In G. M. Garrita (ed), Bergey’s manual of systematic bacteriology, Springer-Verlag, New York, NY vol. 2, part B, 384–401 (2005).
-
- Segura D, Núñez C, Espín G. Azotobacter cysts. eLS Wiley Online Library. 2014
-
- Kennedy C, et al. The nifH, nifM and nifN genes of Azotobacter vinelandii: characterization by Tn5 mutagenesis and isolation from pLARF1 gene banks. Mol. Gen. Genet. 1986;205:318–325. doi: 10.1007/BF00430445. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
