

A single biochemical activity underlies the pleiotropy of the aging-related protein CLK-1
- PMID: 28404998
- PMCID: PMC5429816
- DOI: 10.1038/s41598-017-00754-z
A single biochemical activity underlies the pleiotropy of the aging-related protein CLK-1
Abstract
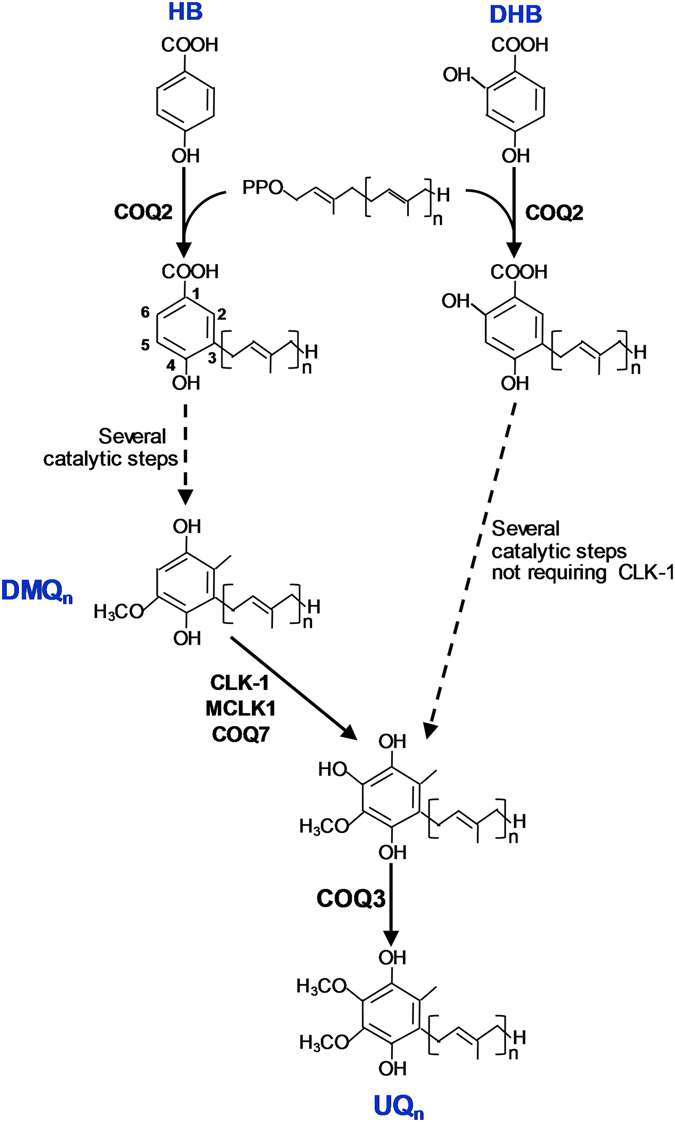
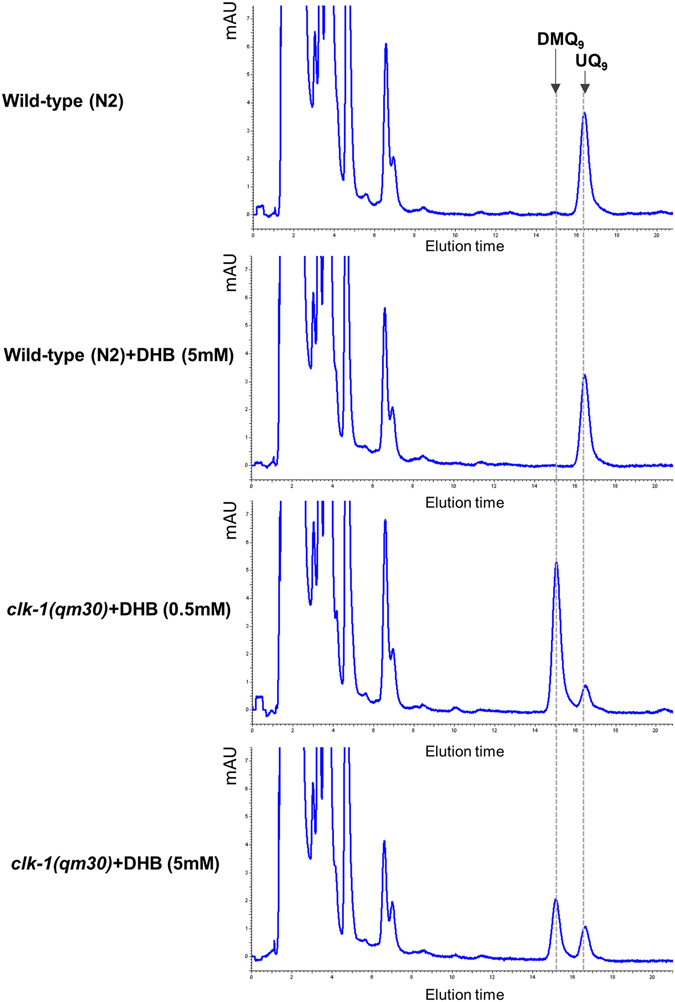
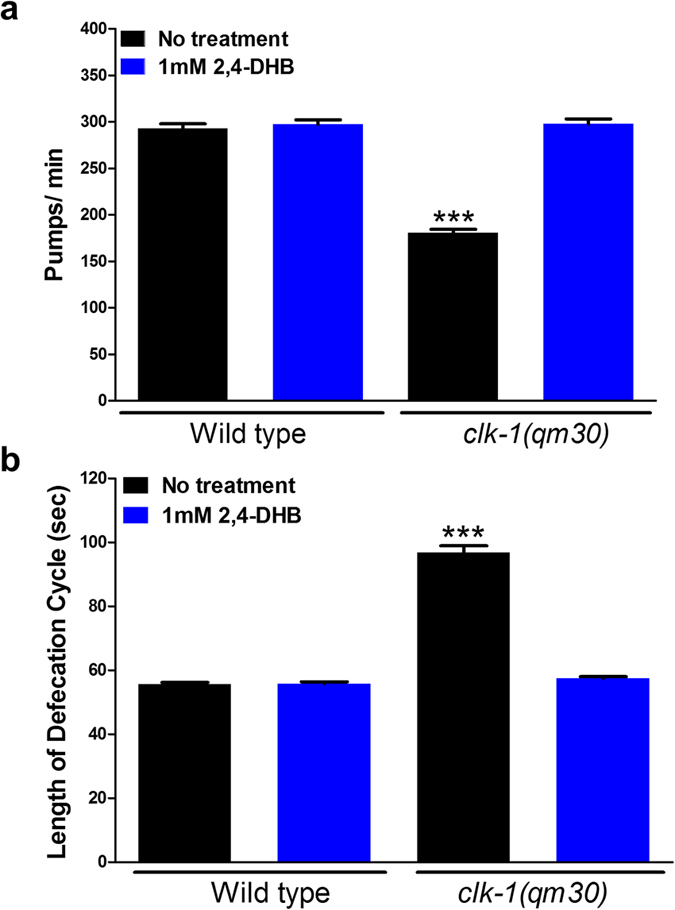
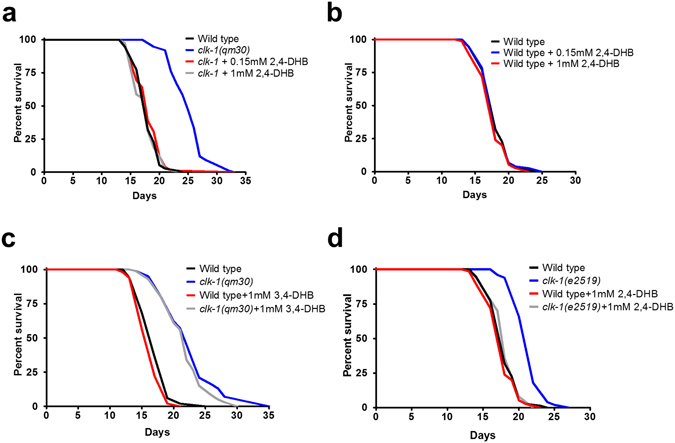
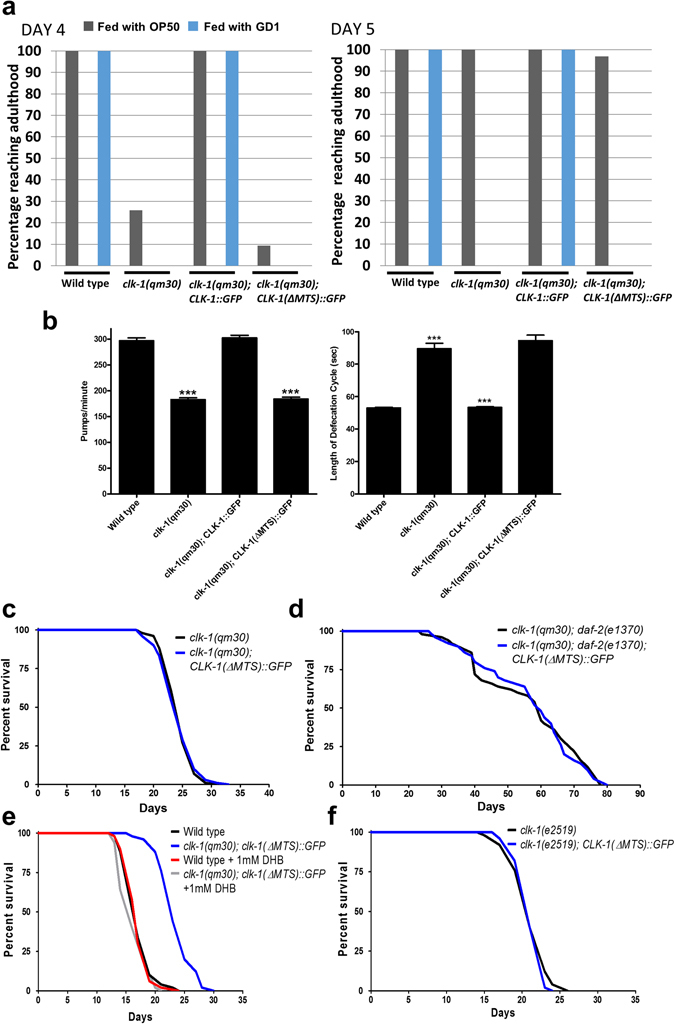
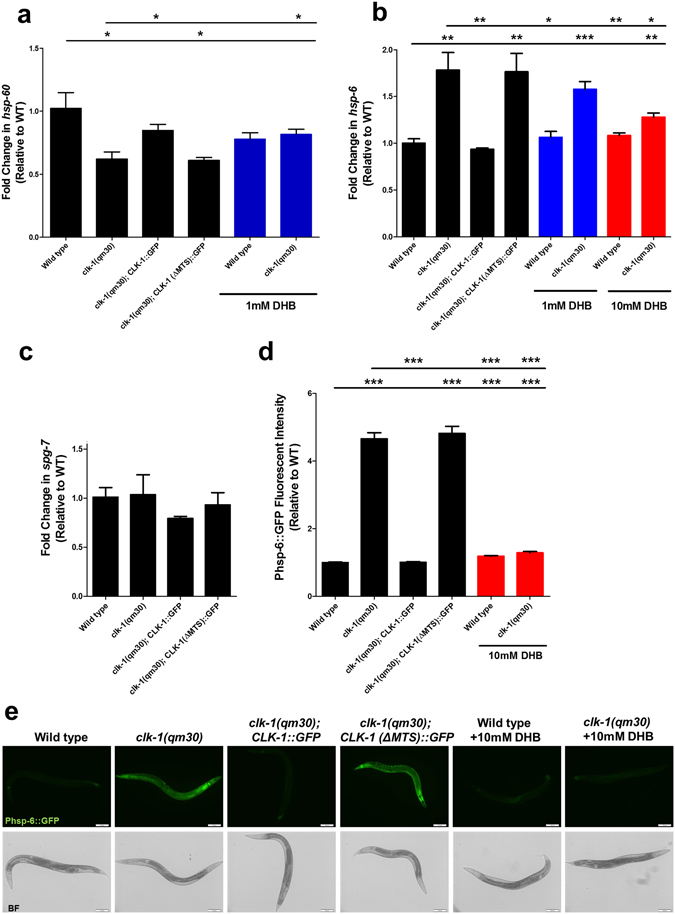
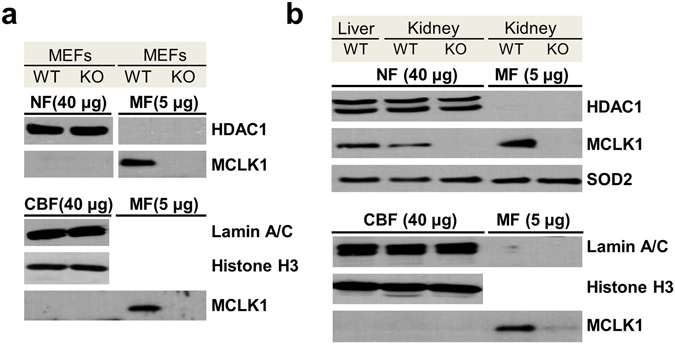
The Caenorhabditis elegans clk-1 gene and the orthologous mouse gene Mclk1 encode a mitochondrial hydroxylase that is necessary for the biosynthesis of ubiquinone (UQ). Mutations in these genes produce broadly pleiotropic phenotypes in both species, including a lengthening of animal lifespan. A number of features of the C. elegans clk-1 mutants, including a maternal effect, particularly extensive pleiotropy, as well as unexplained differences between alleles have suggested that CLK-1/MCLK1 might have additional functions besides that in UQ biosynthesis. In addition, a recent study suggested that a cryptic nuclear localization signal could lead to nuclear localization in cultured mammalian cell lines. Here, by using immunohistochemical techniques in worms and purification techniques in mammalian cells, we failed to detect any nuclear enrichment of the MCLK1 or CLK-1 proteins and any biological activity of a C. elegans CLK-1 protein devoid of a mitochondrial localization sequence. In addition, and most importantly, by pharmacologically restoring UQ biosynthesis in clk-1 null mutants we show that loss of UQ biosynthesis is responsible for all phenotypes resulting from loss of CLK-1, including behavioral phenotypes, altered expression of mitochondrial quality control genes, and lifespan.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
The effect of different ubiquinones on lifespan in Caenorhabditis elegans.Mech Ageing Dev. 2009 Jun;130(6):370-6. doi: 10.1016/j.mad.2009.03.003. Epub 2009 Mar 25. Mech Ageing Dev. 2009. PMID: 19428456 Free PMC article.
-
Genetic and molecular characterization of CLK-1/mCLK1, a conserved determinant of the rate of aging.Exp Gerontol. 2006 Oct;41(10):940-51. doi: 10.1016/j.exger.2006.06.041. Epub 2006 Aug 4. Exp Gerontol. 2006. PMID: 16889924 Review.
-
Altered quinone biosynthesis in the long-lived clk-1 mutants of Caenorhabditis elegans.J Biol Chem. 2001 Mar 16;276(11):7713-6. doi: 10.1074/jbc.C000889200. Epub 2001 Jan 17. J Biol Chem. 2001. PMID: 11244089
-
Uncoupling the pleiotropic phenotypes of clk-1 with tRNA missense suppressors in Caenorhabditis elegans.Mol Cell Biol. 2006 May;26(10):3976-85. doi: 10.1128/MCB.26.10.3976-3985.2006. Mol Cell Biol. 2006. PMID: 16648490 Free PMC article.
-
Quinones in long-lived clk-1 mutants of Caenorhabditis elegans.FEBS Lett. 2002 Feb 13;512(1-3):33-7. doi: 10.1016/s0014-5793(02)02282-2. FEBS Lett. 2002. PMID: 11852047 Review.
Cited by
-
Reduction in the levels of CoQ biosynthetic proteins is related to an increase in lifespan without evidence of hepatic mitohormesis.Sci Rep. 2018 Sep 18;8(1):14013. doi: 10.1038/s41598-018-32190-y. Sci Rep. 2018. PMID: 30228311 Free PMC article.
-
Elevated mitochondrial DNA copy number found in ubiquinone-deficient clk-1 mutants is not rescued by ubiquinone precursor 2-4-dihydroxybenzoate.Mitochondrion. 2021 May;58:38-48. doi: 10.1016/j.mito.2021.02.001. Epub 2021 Feb 11. Mitochondrion. 2021. PMID: 33581333 Free PMC article.
-
Biochemistry of Mitochondrial Coenzyme Q Biosynthesis.Trends Biochem Sci. 2017 Oct;42(10):824-843. doi: 10.1016/j.tibs.2017.06.008. Epub 2017 Sep 17. Trends Biochem Sci. 2017. PMID: 28927698 Free PMC article. Review.
-
A novel COQ7 mutation causing primarily neuromuscular pathology and its treatment options.Mol Genet Metab Rep. 2022 May 5;31:100877. doi: 10.1016/j.ymgmr.2022.100877. eCollection 2022 Jun. Mol Genet Metab Rep. 2022. PMID: 35782625 Free PMC article.
-
Chromatin-remodeling SWI/SNF complex regulates coenzyme Q6 synthesis and a metabolic shift to respiration in yeast.J Biol Chem. 2017 Sep 8;292(36):14851-14866. doi: 10.1074/jbc.M117.798397. Epub 2017 Jul 24. J Biol Chem. 2017. PMID: 28739803 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
