

The chemical basis for electrical signaling
- PMID: 28406893
- PMCID: PMC5464002
- DOI: 10.1038/nchembio.2353
The chemical basis for electrical signaling
Abstract
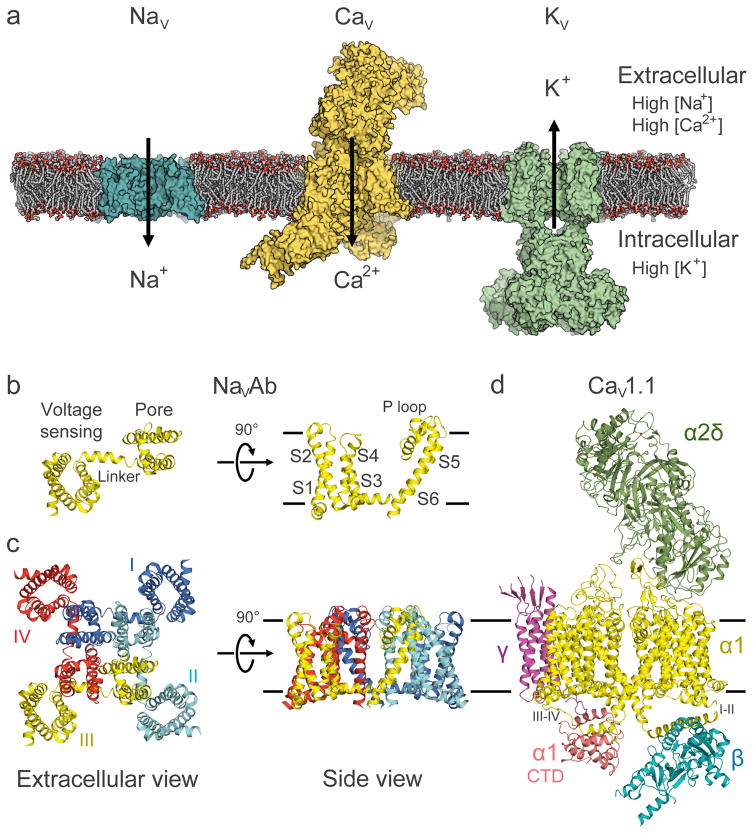
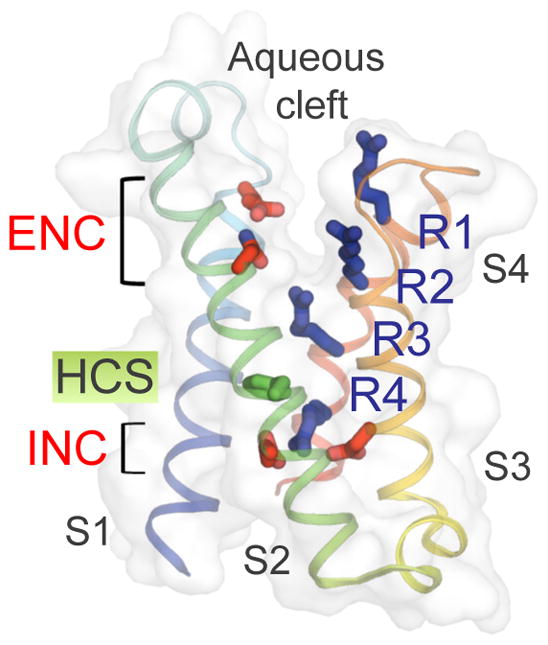
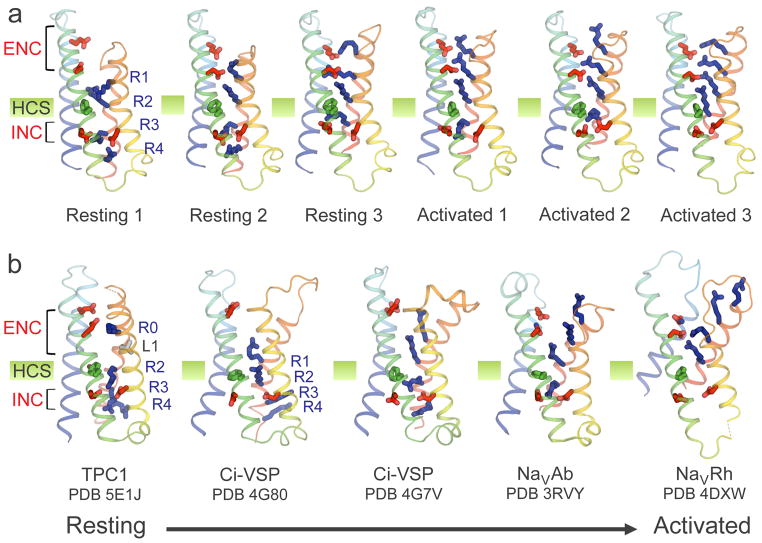
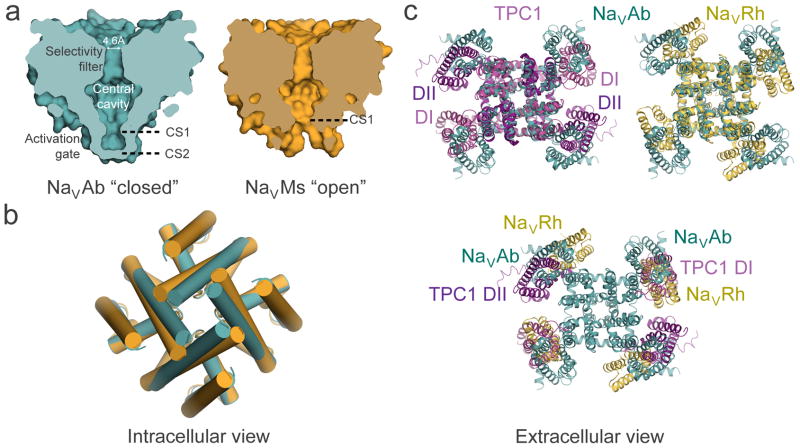
Electrical signals generated by minute currents of ions moving across cell membranes are central to all rapid processes in biology. Initiation and propagation of electrical signals requires voltage-gated sodium (NaV) and calcium (CaV) channels. These channels contain a tetramer of membrane-bound subunits or domains comprising a voltage sensor and a pore module. Voltage-dependent activation occurs as membrane depolarization drives outward movements of positive gating changes in the voltage sensor via a sliding-helix mechanism, which leads to a conformational change in the pore module that opens its intracellular activation gate. A unique negatively charged site in the selectivity filter conducts hydrated Na+ or Ca2+ rapidly and selectively. Ion conductance is terminated by voltage-dependent inactivation, which causes asymmetric pore collapse. This Review focuses on recent advances in structure and function of NaV and CaV channels that expand our current understanding of the chemical basis for electrical signaling mechanisms conserved from bacteria to humans.
Figures
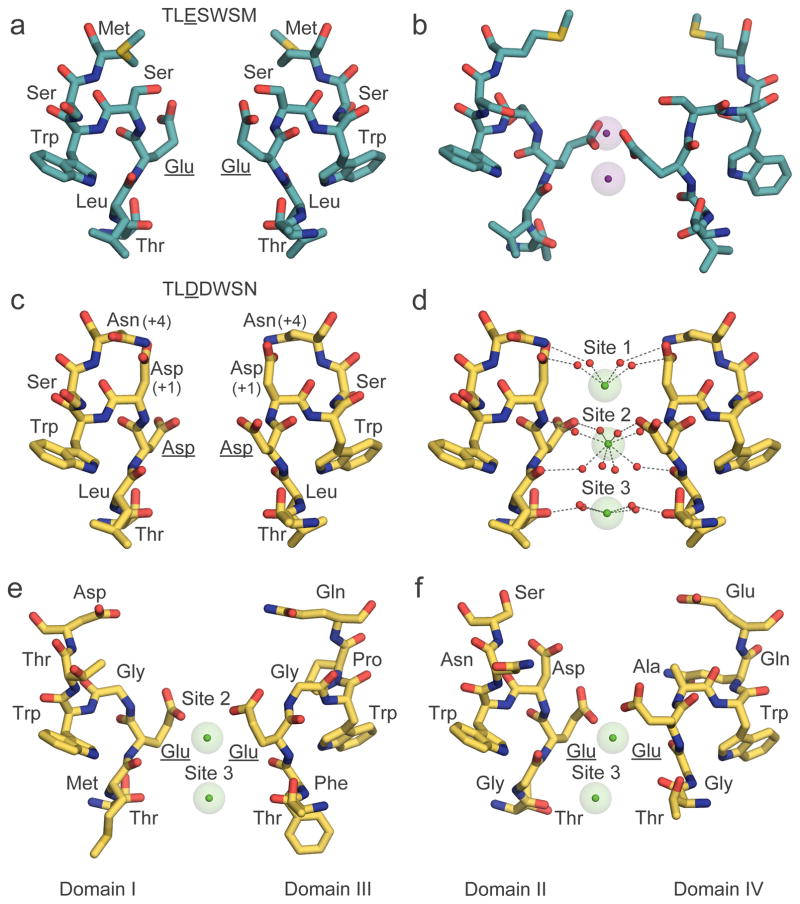
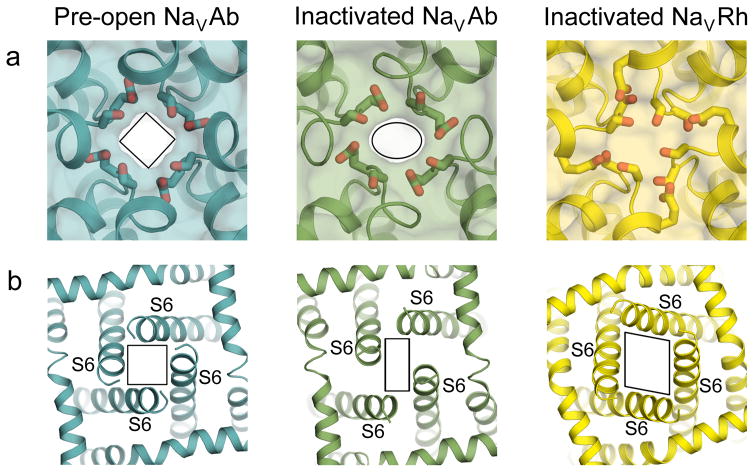
References
-
- Hille B. Ionic Channels of Excitable Membranes. 3. Sinauer Associates Inc; 2001.
-
- Long SB, Campbell EB, Mackinnon R. Crystal structure of a mammalian voltage-dependent Shaker family K+ channel. Science. 2005;309:897–903. - PubMed
References*
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
