

Transcriptional Elongation of HSV Immediate Early Genes by the Super Elongation Complex Drives Lytic Infection and Reactivation from Latency
- PMID: 28407486
- PMCID: PMC5997188
- DOI: 10.1016/j.chom.2017.03.007
Transcriptional Elongation of HSV Immediate Early Genes by the Super Elongation Complex Drives Lytic Infection and Reactivation from Latency
Abstract
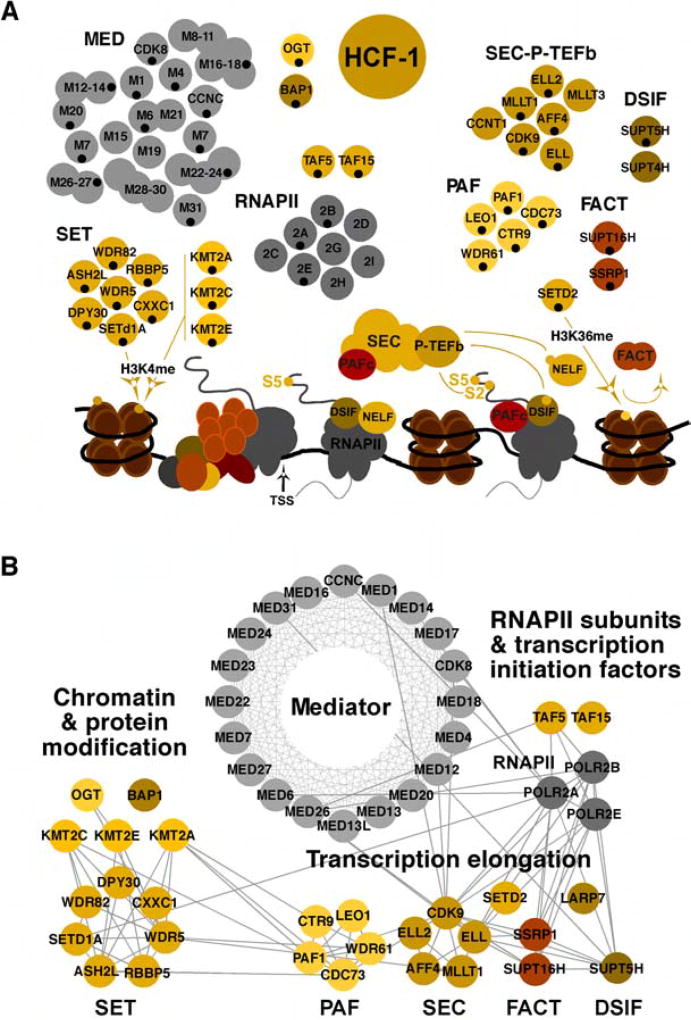
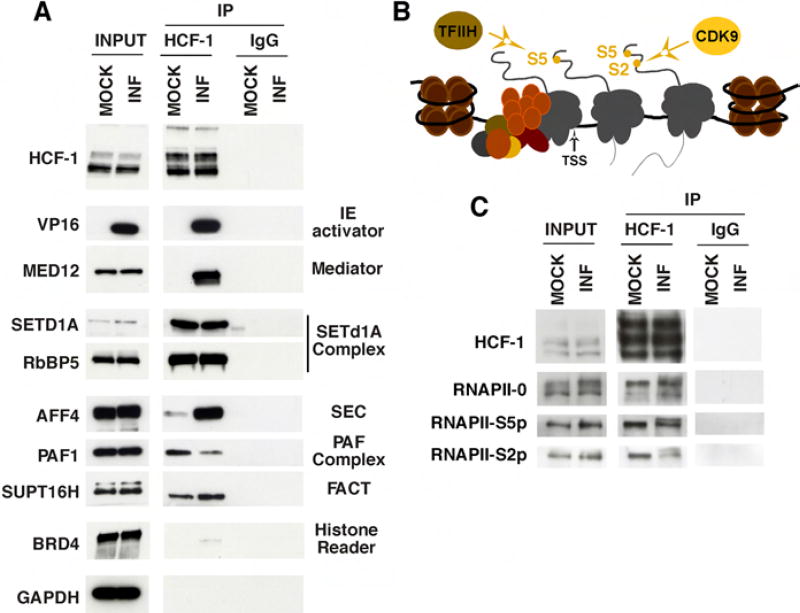
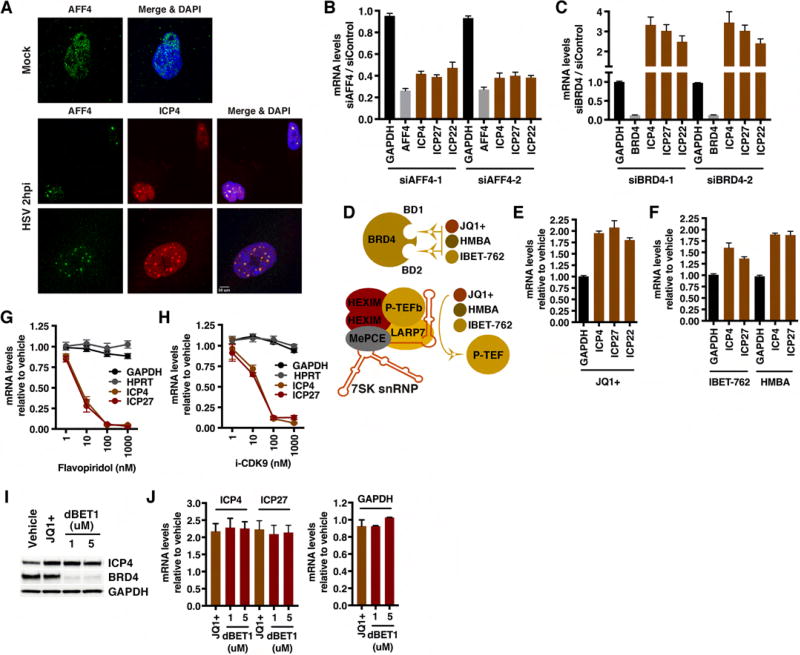
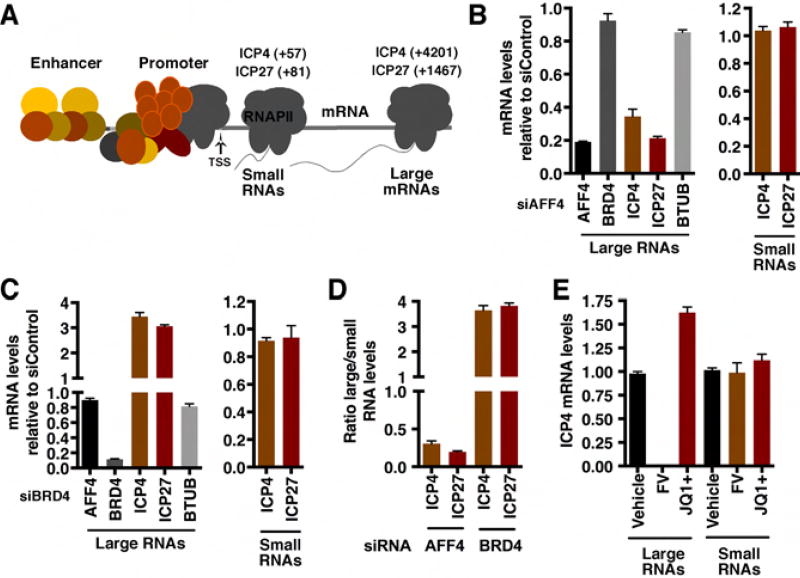
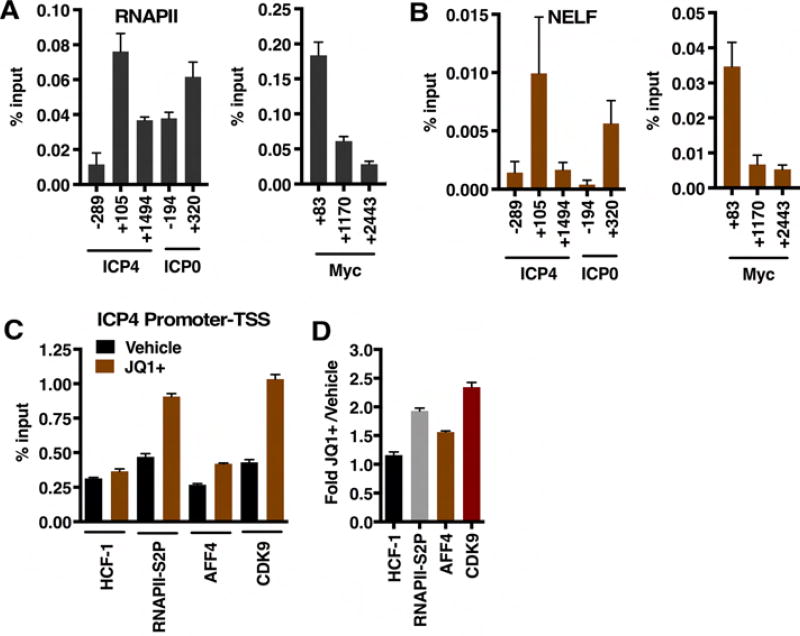
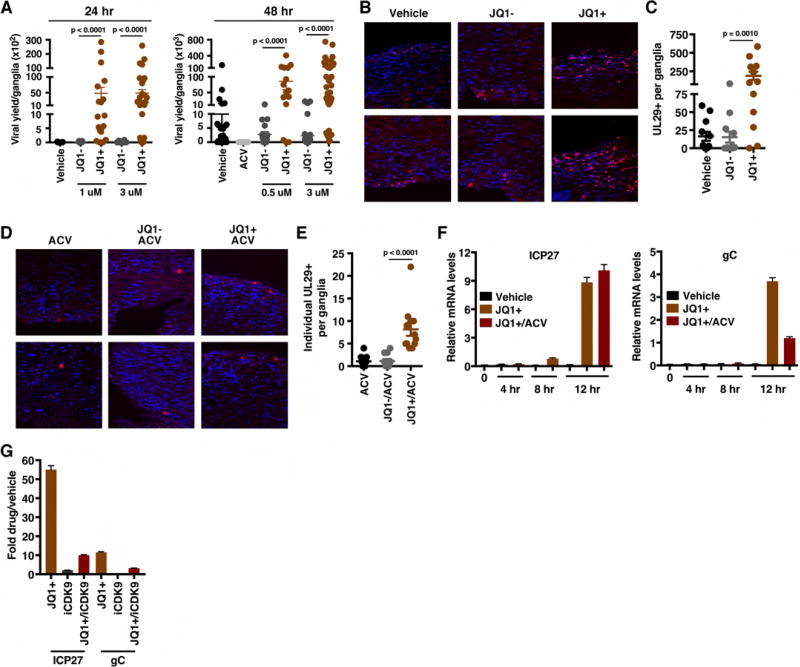
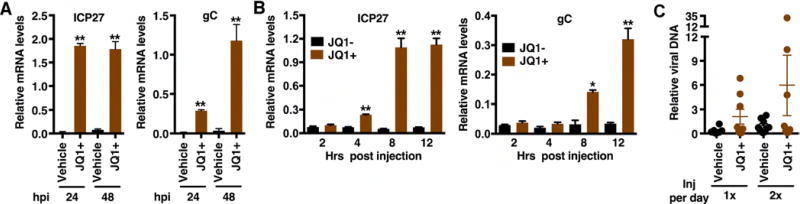
The cellular transcriptional coactivator HCF-1 is required for initiation of herpes simplex virus (HSV) lytic infection and for reactivation from latency in sensory neurons. HCF-1 stabilizes the viral Immediate Early (IE) gene enhancer complex and mediates chromatin transitions to promote IE transcription initiation. In infected cells, HCF-1 was also found to be associated with a network of transcription elongation components including the super elongation complex (SEC). IE genes exhibit characteristics of genes controlled by transcriptional elongation, and the SEC-P-TEFb complex is specifically required to drive the levels of productive IE mRNAs. Significantly, compounds that enhance the levels of SEC-P-TEFb also potently stimulated HSV reactivation from latency both in a sensory ganglia model system and in vivo. Thus, transcriptional elongation of HSV IE genes is a key limiting parameter governing both the initiation of HSV infection and reactivation of latent genomes.
Keywords: P-TEFb; herpes simplex virus; host cell factor-1; latency; super elongation complex; transcriptional elongation.
Published by Elsevier Inc.
Figures
References
-
- Bartholomeeusen K, Xiang Y, Fujinaga K, Peterlin BM. Bromodomain and extra-terminal (BET) bromodomain inhibition activate transcription via transient release of positive transcription elongation factor b (P-TEFb) from 7SK small nuclear ribonucleoprotein. J Biol Chem. 2012;287:36609–36616. - PMC - PubMed
-
- Bernstein DI, Kappes JC. Enhanced in vitro reactivation of latent herpes simplex virus from neural and peripheral tissues with hexamethylenebisacetamide. Arch Virol. 1988;99:57–65. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
