

Human-derived gut microbiota modulates colonic secretion in mice by regulating 5-HT3 receptor expression via acetate production
- PMID: 28408644
- PMCID: PMC5538830
- DOI: 10.1152/ajpgi.00448.2016
Human-derived gut microbiota modulates colonic secretion in mice by regulating 5-HT3 receptor expression via acetate production
Abstract
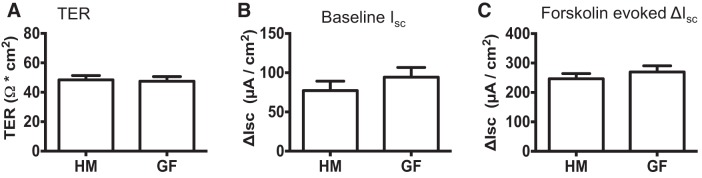
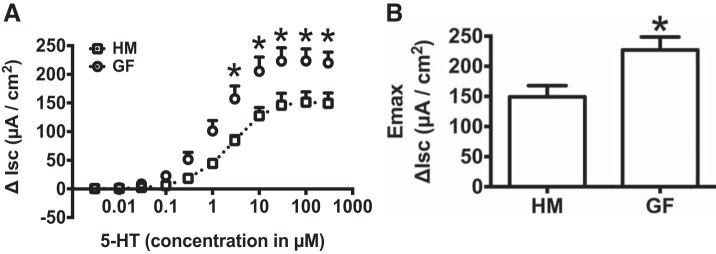
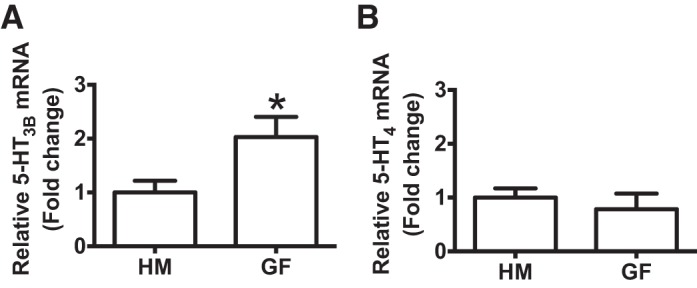
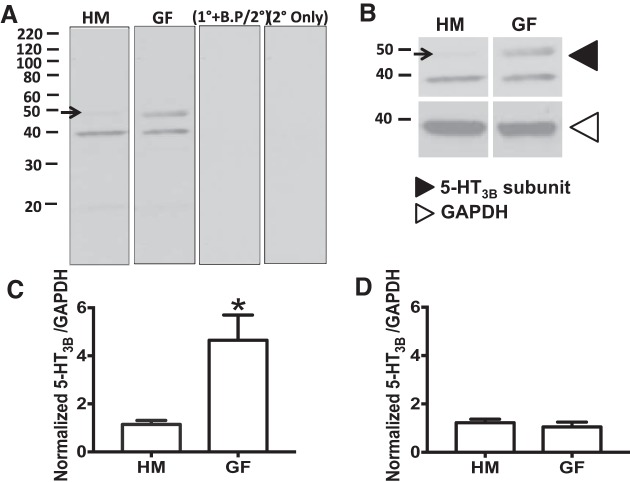
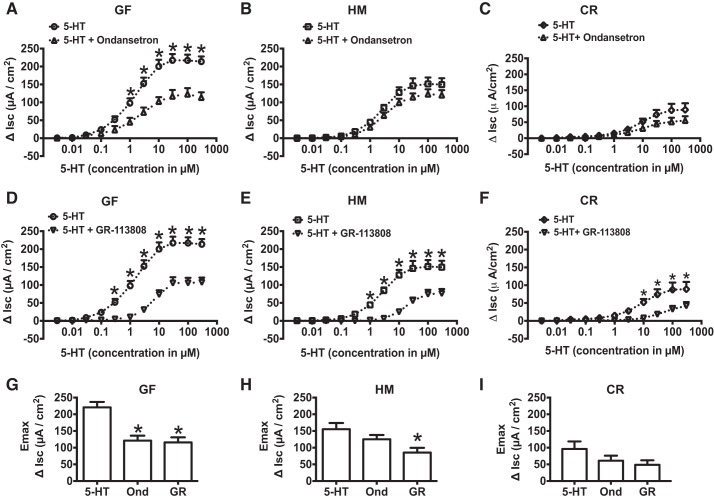
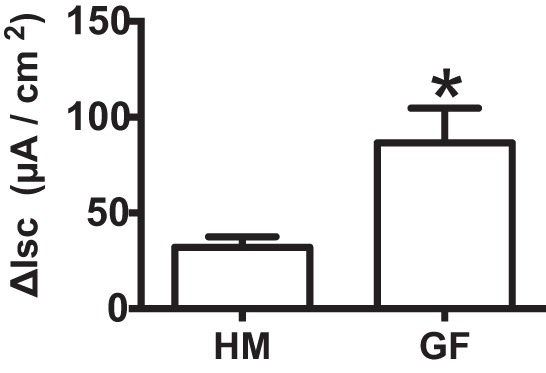
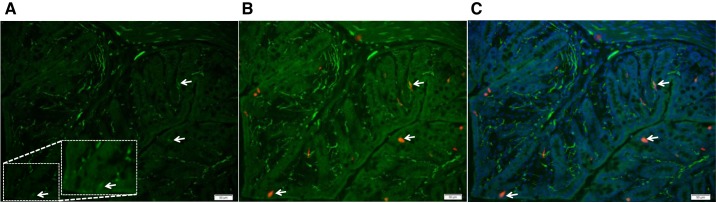
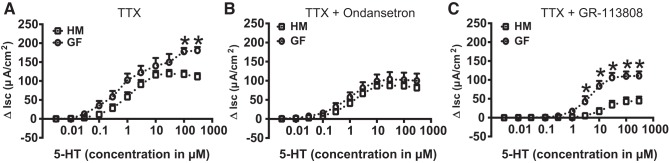
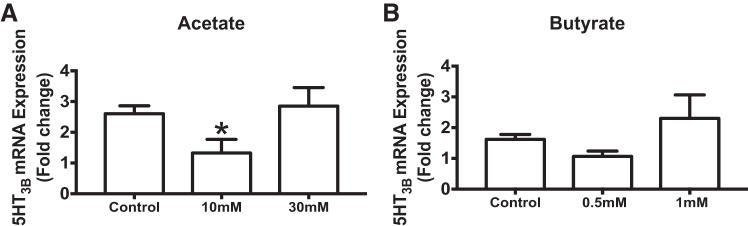
Serotonin [5-hydroxytryptamine (5-HT)], an important neurotransmitter and a paracrine messenger in the gastrointestinal tract, regulates intestinal secretion by its action primarily on 5-HT3 and 5-HT4 receptors. Recent studies highlight the role of gut microbiota in 5-HT biosynthesis. In this study, we determine whether human-derived gut microbiota affects host secretory response to 5-HT and 5-HT receptor expression. We used proximal colonic mucosa-submucosa preparation from age-matched Swiss Webster germ-free (GF) and humanized (HM; ex-GF colonized with human gut microbiota) mice. 5-HT evoked a significantly greater increase in short-circuit current (ΔIsc) in GF compared with HM mice. Additionally, 5-HT3 receptor mRNA and protein expression was significantly higher in GF compared with HM mice. Ondansetron, a 5-HT3 receptor antagonist, inhibited 5-HT-evoked ΔIsc in GF mice but not in HM mice. Furthermore, a 5-HT3 receptor-selective agonist, 2-methyl-5-hydroxytryptamine hydrochloride, evoked a significantly higher ΔIsc in GF compared with HM mice. Immunohistochemistry in 5-HT3A-green fluorescent protein mice localized 5-HT3 receptor expression to enterochromaffin cells in addition to nerve fibers. The significant difference in 5-HT-evoked ΔIsc between GF and HM mice persisted in the presence of tetrodotoxin (TTX) but was lost after ondansetron application in the presence of TTX. Application of acetate (10 mM) significantly lowered 5-HT3 receptor mRNA in GF mouse colonoids. We conclude that host secretory response to 5-HT may be modulated by gut microbiota regulation of 5-HT3 receptor expression via acetate production. Epithelial 5-HT3 receptor may function as a mediator of gut microbiota-driven change in intestinal secretion.NEW & NOTEWORTHY We found that gut microbiota alters serotonin (5-HT)-evoked intestinal secretion in a 5-HT3 receptor-dependent mechanism and gut microbiota metabolite acetate alters 5-HT3 receptor expression in colonoids.View this article's corresponding video summary at https://www.youtube.com/watch?v=aOMYJMuLTcw&feature=youtu.be.
Keywords: 5-hydroxytryptamine type 3; irritable bowel syndrome; microbiome; motility; physiology; secretion.
Copyright © 2017 the American Physiological Society.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases