

Karyotype diversity and chromosomal organization of repetitive DNA in Tityus obscurus (Scorpiones, Buthidae)
- PMID: 28412934
- PMCID: PMC5392961
- DOI: 10.1186/s12863-017-0494-6
Karyotype diversity and chromosomal organization of repetitive DNA in Tityus obscurus (Scorpiones, Buthidae)
Abstract
Background: Holocentric chromosomes occur in approximately 750 species of eukaryotes. Among them, the genus Tityus (Scorpiones, Buthidae) has a labile karyotype that shows complex multivalent associations during male meiosis. Thus, taking advantage of the excellent model provided by the Buthidae scorpions, here we analyzed the chromosomal distribution of several repetitive DNA classes on the holocentric chromosomes of different populations of the species Tityus obscurus Gervais, 1843, highlighting their involvement in the karyotypic differences found among them.
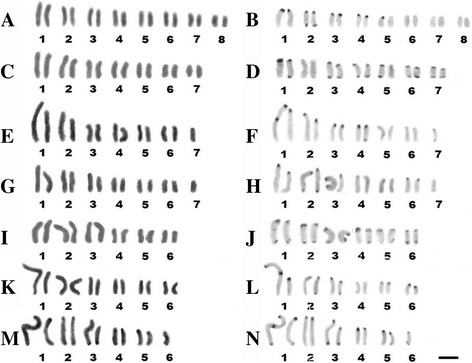
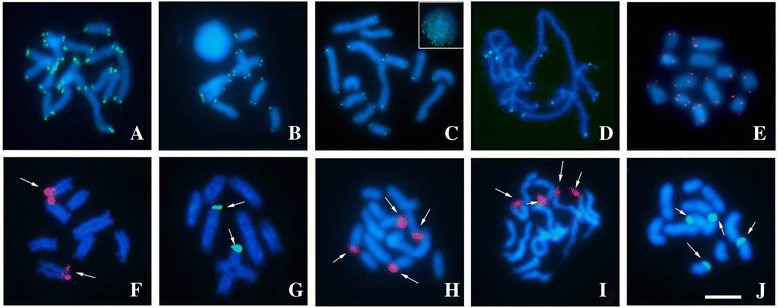
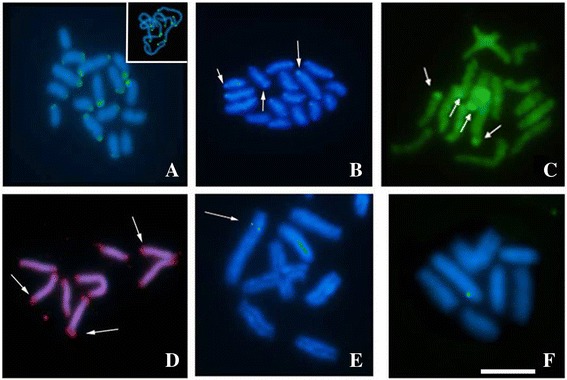
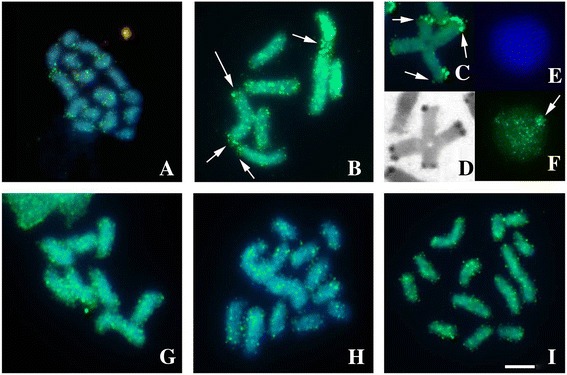
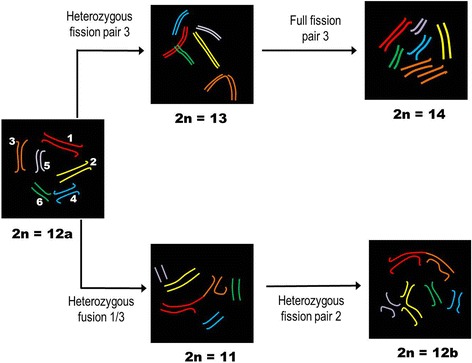
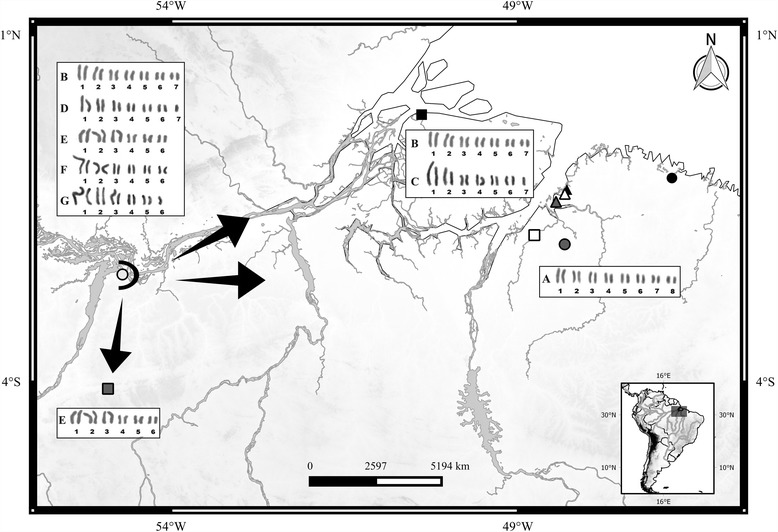
Results: This species shows inter- and intrapopulational karyotype variation, with seven distinct cytotypes: A (2n = 16), B (2n = 14), C (2n = 13), D (2n = 13), E (2n = 12), F (2n = 12) and G (2n = 11). Furthermore, exhibits achiasmatic male meiosis and lacks heteromorphic sex chromosomes. Trivalent and quadrivalent meiotic associations were found in some cytotypes. In them, 45S rDNAs were found in the terminal portions of two pairs, while TTAGG repeats were found only at the end of the chromosomes. In the cytotype A (2n = 16), the U2 snRNA gene mapped to pair 1, while the H3 histone cluster and C 0 t-1 DNA fraction was terminally distributed on all pairs. Mariner transposons were found throughout the chromosomes, with the exception of one individual of cytotype A (2n = 16), in which it was concentrated in heterochromatic regions.
Conclusions: Chromosomal variability found in T. obscurus are due to rearrangements of the type fusion/fission and reciprocal translocations in heterozygous. These karyotype differences follow a geographical pattern and may be contributing to reproductive isolation between populations analyzed. Our results also demonstrate high mobility of histone H3 genes. In contrast, other multigene families (45S rDNA and U2 snRNA) have conserved distribution among individuals. The accumulation of repetitive sequences in distal regions of T. obscurus chromosomes, suggests that end of chromosome are not covered by the kinetochore.
Keywords: Holocentric chromosomes; Multivalent association; Repetitive DNA; Scorpiones; Tityus.
Figures

Similar articles
-
Location of 45S Ribosomal Genes in Mitotic and Meiotic Chromosomes of Buthid Scorpions.Zoolog Sci. 2014 Sep;31(9):603-7. doi: 10.2108/zs140005. Zoolog Sci. 2014. PMID: 25186932
-
Male and female meiosis in the mountain scorpion Zabius fuscus (Scorpiones, Buthidae): heterochromatin, rDNA and TTAGG telomeric repeats.Genetica. 2015 Aug;143(4):393-401. doi: 10.1007/s10709-015-9838-1. Epub 2015 Apr 22. Genetica. 2015. PMID: 25899730
-
Insights into the origin of the high variability of multivalent-meiotic associations in holocentric chromosomes of Tityus (Archaeotityus) scorpions.PLoS One. 2018 Feb 21;13(2):e0192070. doi: 10.1371/journal.pone.0192070. eCollection 2018. PLoS One. 2018. PMID: 29466400 Free PMC article.
-
The karyotypes and evolution of ZZ/ZW sex chromosomes in the genus Characidium (Characiformes, Crenuchidae).Comp Cytogenet. 2018 Oct 2;12(3):421-438. doi: 10.3897/CompCytogen.v12i3.28736. eCollection 2018. Comp Cytogenet. 2018. PMID: 30310546 Free PMC article. Review.
-
Mosquito genomes: structure, organization, and evolution.Adv Genet. 1999;41:1-33. doi: 10.1016/s0065-2660(08)60149-2. Adv Genet. 1999. PMID: 10494615 Review.
Cited by
-
Advances in Understanding the Karyotype Evolution of Tetrapulmonata and Two Other Arachnid Taxa, Ricinulei and Solifugae.Genes (Basel). 2025 Feb 8;16(2):207. doi: 10.3390/genes16020207. Genes (Basel). 2025. PMID: 40004536 Free PMC article.
-
Insights into the Karyotype Evolution of Charinidae, the Early-Diverging Clade of Whip Spiders (Arachnida: Amblypygi).Animals (Basel). 2021 Nov 12;11(11):3233. doi: 10.3390/ani11113233. Animals (Basel). 2021. PMID: 34827965 Free PMC article.
-
Chromosome diversity in Buthidae and Chactidae scorpions from Brazilian fauna: Diploid number and distribution of repetitive DNA sequences.Genet Mol Biol. 2023 May 15;46(2):e20220083. doi: 10.1590/1678-4685-GMB-2022-0083. eCollection 2023. Genet Mol Biol. 2023. PMID: 37216321 Free PMC article.
-
Kinetic Activity of Chromosomes and Expression of Recombination Genes in Achiasmatic Meiosis of Tityus (Archaeotityus) Scorpions.Int J Mol Sci. 2022 Aug 16;23(16):9179. doi: 10.3390/ijms23169179. Int J Mol Sci. 2022. PMID: 36012447 Free PMC article.
-
Meiosis in the scorpion Tityus silvestris: new insights into achiasmatic chromosomes.Biol Open. 2019 May 9;8(5):bio040352. doi: 10.1242/bio.040352. Biol Open. 2019. PMID: 31072909 Free PMC article.
References
-
- Chung KS, Weber JA, Hipp AL. Dynamics of chromosome number and genome size variation in acyogeneically variable sedge (Carex scoparia var, scoparia, Cyperaceae) Am J Bot. 2011;98:129–122. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
