

Structure and function of Helicobacter pylori CagA, the first-identified bacterial protein involved in human cancer
- PMID: 28413197
- PMCID: PMC5489429
- DOI: 10.2183/pjab.93.013
Structure and function of Helicobacter pylori CagA, the first-identified bacterial protein involved in human cancer
Abstract
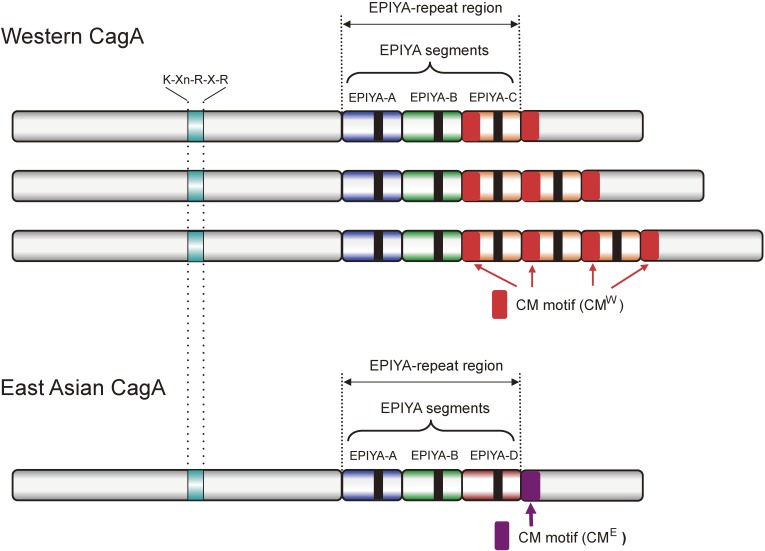
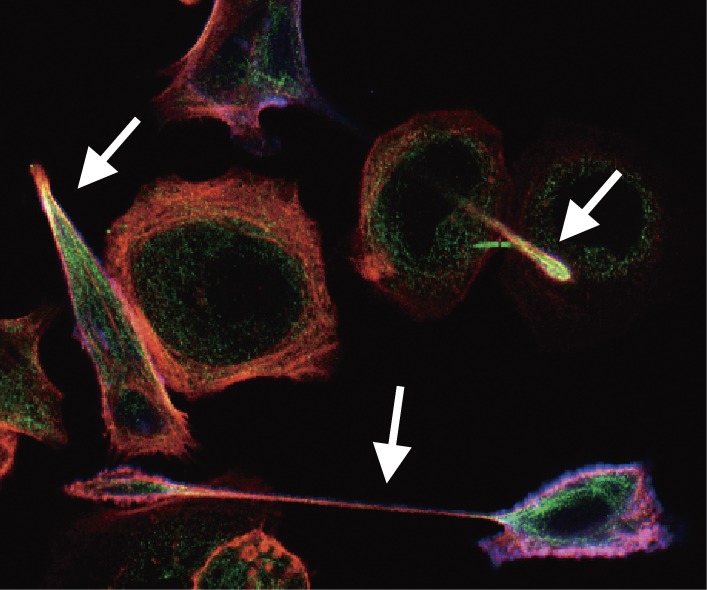
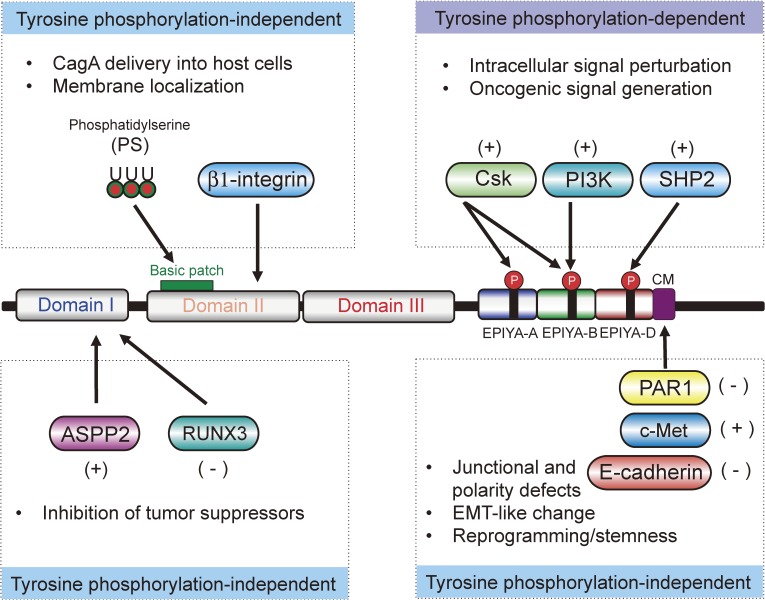
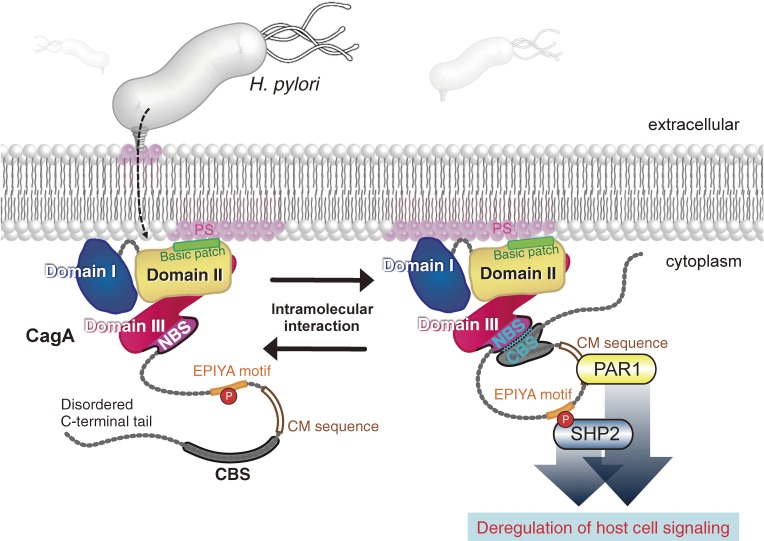
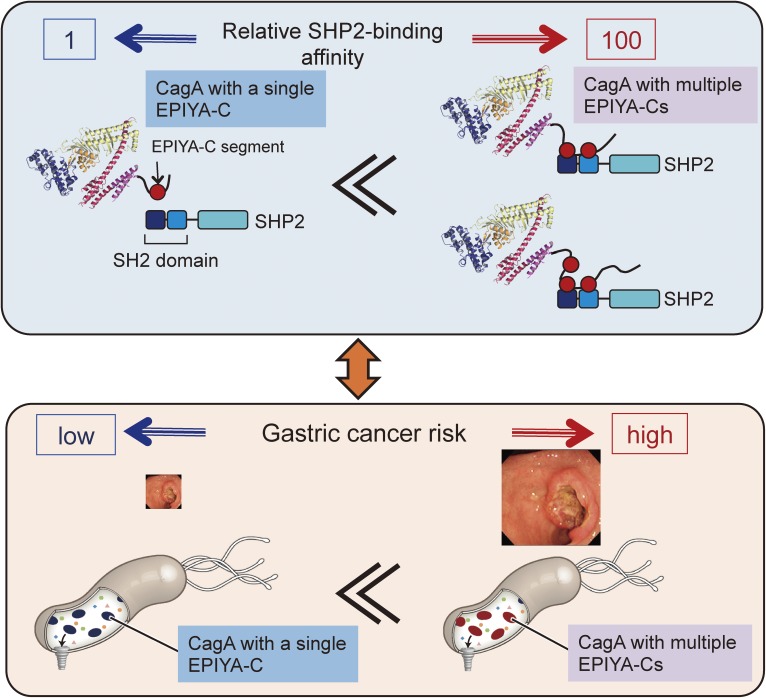
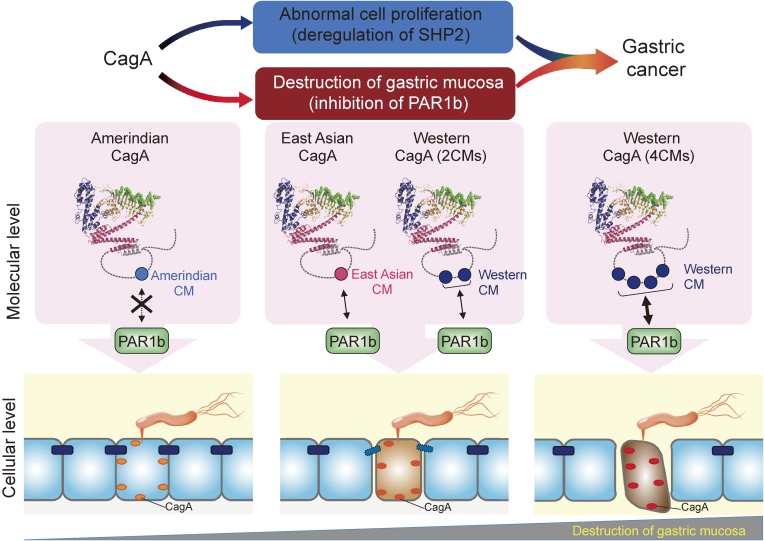
Chronic infection with Helicobacter pylori cagA-positive strains is the strongest risk factor of gastric cancer. The cagA gene-encoded CagA protein is delivered into gastric epithelial cells via bacterial type IV secretion, where it undergoes tyrosine phosphorylation at the Glu-Pro-Ile-Tyr-Ala (EPIYA) motifs. Delivered CagA then acts as a non-physiological scaffold/hub protein by interacting with multiple host signaling molecules, most notably the pro-oncogenic phosphatase SHP2 and the polarity-regulating kinase PAR1/MARK, in both tyrosine phosphorylation-dependent and -independent manners. CagA-mediated manipulation of intracellular signaling promotes neoplastic transformation of gastric epithelial cells. Transgenic expression of CagA in experimental animals has confirmed the oncogenic potential of the bacterial protein. Structural polymorphism of CagA influences its scaffold function, which may underlie the geographic difference in the incidence of gastric cancer. Since CagA is no longer required for the maintenance of established gastric cancer cells, studying the role of CagA during neoplastic transformation will provide an excellent opportunity to understand molecular processes underlying "Hit-and-Run" carcinogenesis.
Figures

References
-
- Ferlay J., Soerjomataram I., Dikshit R., Eser S., Mathers C., Rebelo M., Parkin D.M., Forman D., Bray F. (2015) Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 136, E359–E386. - PubMed
-
- Lauren P. (1965) The two histological main types of gastric carcinoma: Diffuse and so-called intestinal-type carcinoma. An attempt at a histo-clinical classification. Acta Pathol. Microbiol. Scand. 64, 31–49. - PubMed
-
- Correa P. (1992) Human gastric carcinogenesis: a multistep and multifactorial process — first American Cancer Society award lecture on cancer epidemiology and prevention. Cancer Res. 52, 6735–6740. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources