

Alternatively activated macrophages do not synthesize catecholamines or contribute to adipose tissue adaptive thermogenesis
- PMID: 28414329
- PMCID: PMC5420449
- DOI: 10.1038/nm.4316
Alternatively activated macrophages do not synthesize catecholamines or contribute to adipose tissue adaptive thermogenesis
Abstract
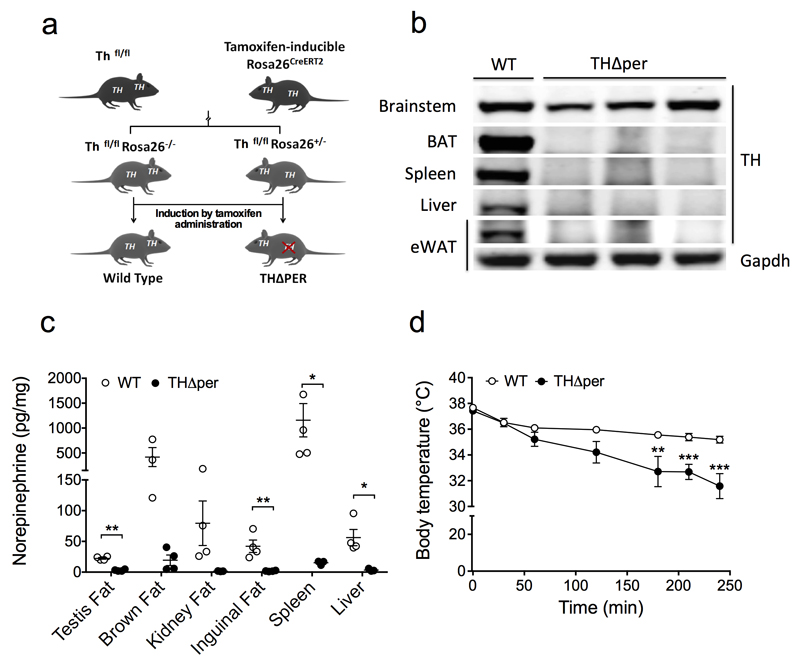
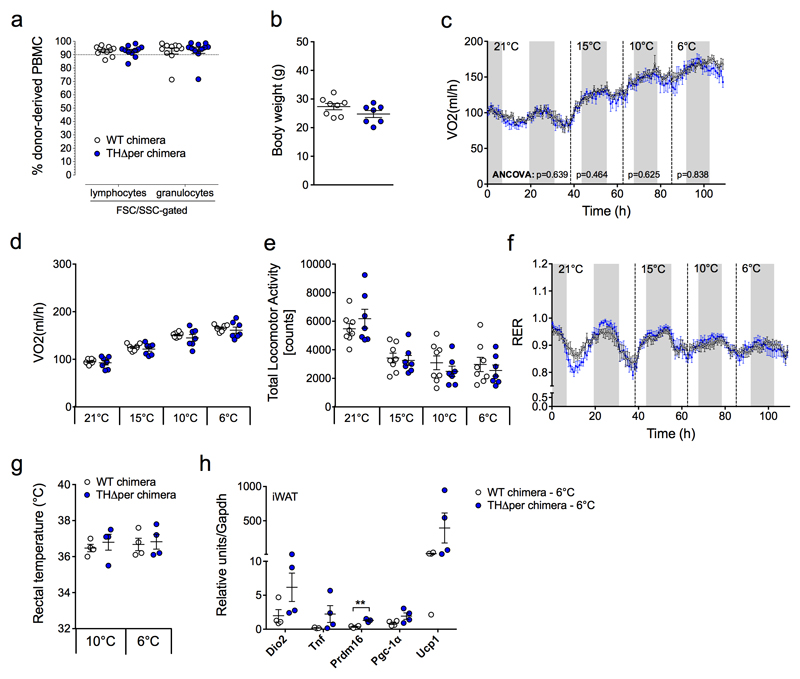
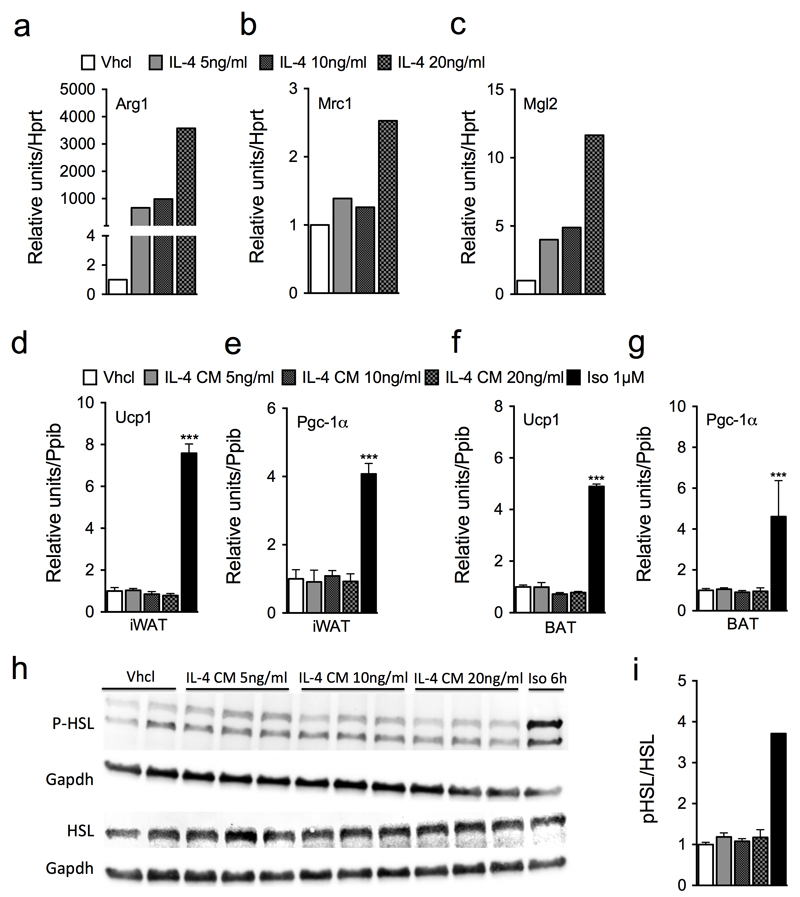
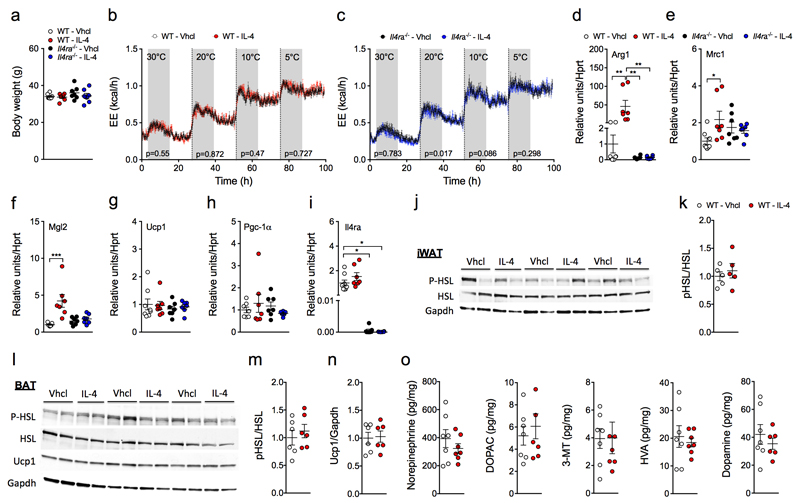
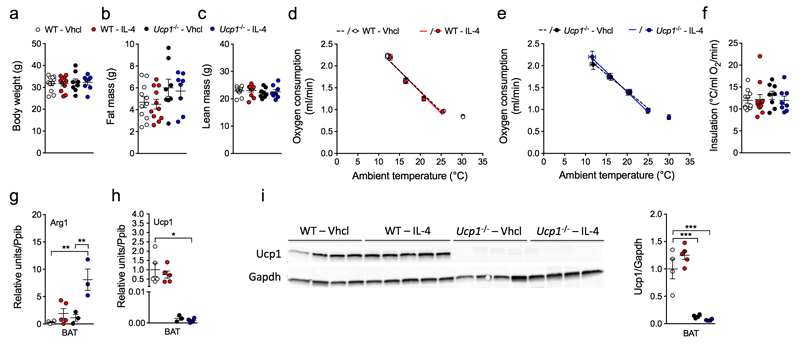
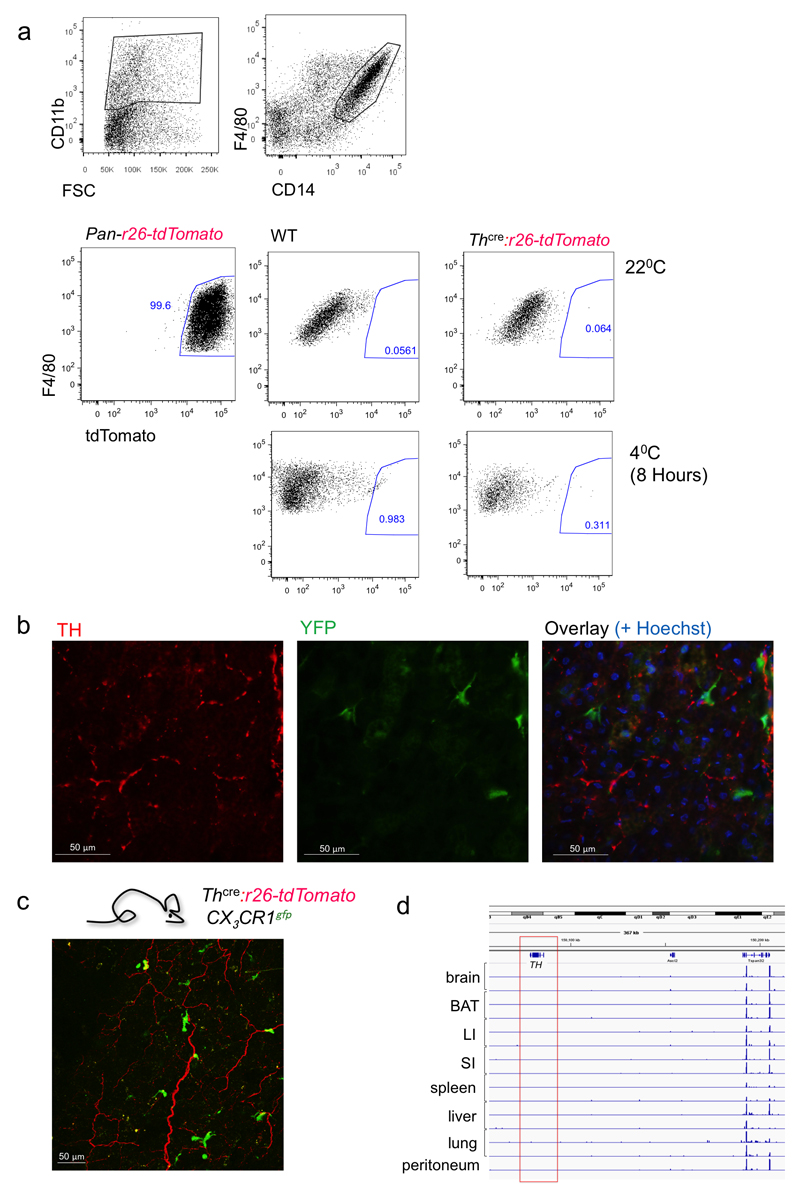
Adaptive thermogenesis is the process of heat generation in response to cold stimulation. It is under the control of the sympathetic nervous system, whose chief effector is the catecholamine norepinephrine (NE). NE enhances thermogenesis through β3-adrenergic receptors to activate brown adipose tissue and by 'browning' white adipose tissue. Recent studies have reported that alternative activation of macrophages in response to interleukin (IL)-4 stimulation induces the expression of tyrosine hydroxylase (TH), a key enzyme in the catecholamine synthesis pathway, and that this activation provides an alternative source of locally produced catecholamines during the thermogenic process. Here we report that the deletion of Th in hematopoietic cells of adult mice neither alters energy expenditure upon cold exposure nor reduces browning in inguinal adipose tissue. Bone marrow-derived macrophages did not release NE in response to stimulation with IL-4, and conditioned media from IL-4-stimulated macrophages failed to induce expression of thermogenic genes, such as uncoupling protein 1 (Ucp1), in adipocytes cultured with the conditioned media. Furthermore, chronic treatment with IL-4 failed to increase energy expenditure in wild-type, Ucp1-/- and interleukin-4 receptor-α double-negative (Il4ra-/-) mice. In agreement with these findings, adipose-tissue-resident macrophages did not express TH. Thus, we conclude that alternatively activated macrophages do not synthesize relevant amounts of catecholamines, and hence, are not likely to have a direct role in adipocyte metabolism or adaptive thermogenesis.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Regulation of Brown and White Adipocyte Transcriptome by the Transcriptional Coactivator NT-PGC-1α.PLoS One. 2016 Jul 25;11(7):e0159990. doi: 10.1371/journal.pone.0159990. eCollection 2016. PLoS One. 2016. PMID: 27454177 Free PMC article.
-
Alternatively activated macrophages produce catecholamines to sustain adaptive thermogenesis.Nature. 2011 Nov 20;480(7375):104-8. doi: 10.1038/nature10653. Nature. 2011. PMID: 22101429 Free PMC article.
-
Continuous cold exposure induces an anti-inflammatory response in mesenteric adipose tissue associated with catecholamine production and thermogenin expression in rats.Endocr Regul. 2016 Jul;50(3):137-44. doi: 10.1515/enr-2016-0015. Endocr Regul. 2016. PMID: 27560796
-
Thermogenesis in Adipose Tissue Activated by Thyroid Hormone.Int J Mol Sci. 2020 Apr 24;21(8):3020. doi: 10.3390/ijms21083020. Int J Mol Sci. 2020. PMID: 32344721 Free PMC article. Review.
-
Brown adipose tissue: Updates in cellular and molecular biology.Tissue Cell. 2016 Oct;48(5):452-60. doi: 10.1016/j.tice.2016.08.001. Epub 2016 Aug 8. Tissue Cell. 2016. PMID: 27561621 Review.
Cited by
-
GIPR Signaling in Immune Cells Maintains Metabolically Beneficial Type 2 Immune Responses in the White Fat From Obese Mice.Front Immunol. 2021 Feb 25;12:643144. doi: 10.3389/fimmu.2021.643144. eCollection 2021. Front Immunol. 2021. PMID: 33717200 Free PMC article.
-
Food for thought - ILC metabolism in the context of helminth infections.Mucosal Immunol. 2022 Jun;15(6):1234-1242. doi: 10.1038/s41385-022-00559-y. Epub 2022 Aug 31. Mucosal Immunol. 2022. PMID: 36045216 Free PMC article. Review.
-
Remodeling of Adipose Tissues by Fatty Acids: Mechanistic Update on Browning and Thermogenesis by n-3 Polyunsaturated Fatty Acids.Pharm Res. 2023 Feb;40(2):467-480. doi: 10.1007/s11095-022-03377-w. Epub 2022 Sep 1. Pharm Res. 2023. PMID: 36050546 Review.
-
A Renewable Source of Human Beige Adipocytes for Development of Therapies to Treat Metabolic Syndrome.Cell Rep. 2018 Dec 11;25(11):3215-3228.e9. doi: 10.1016/j.celrep.2018.11.037. Cell Rep. 2018. PMID: 30540952 Free PMC article.
-
Glucocorticoid/Adiponectin Axis Mediates Full Activation of Cold-Induced Beige Fat Thermogenesis.Biomolecules. 2021 Oct 23;11(11):1573. doi: 10.3390/biom11111573. Biomolecules. 2021. PMID: 34827571 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
