

Neuronal cell fate specification by the molecular convergence of different spatio-temporal cues on a common initiator terminal selector gene
- PMID: 28414802
- PMCID: PMC5411104
- DOI: 10.1371/journal.pgen.1006729
Neuronal cell fate specification by the molecular convergence of different spatio-temporal cues on a common initiator terminal selector gene
Abstract
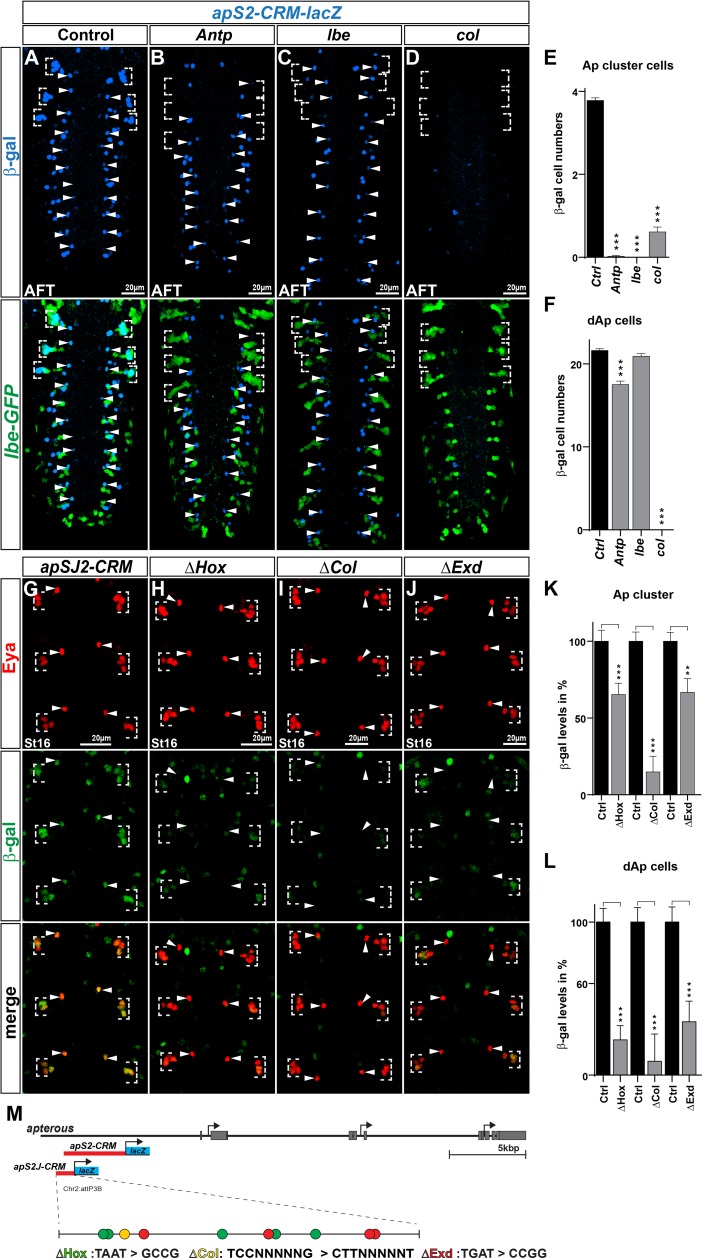
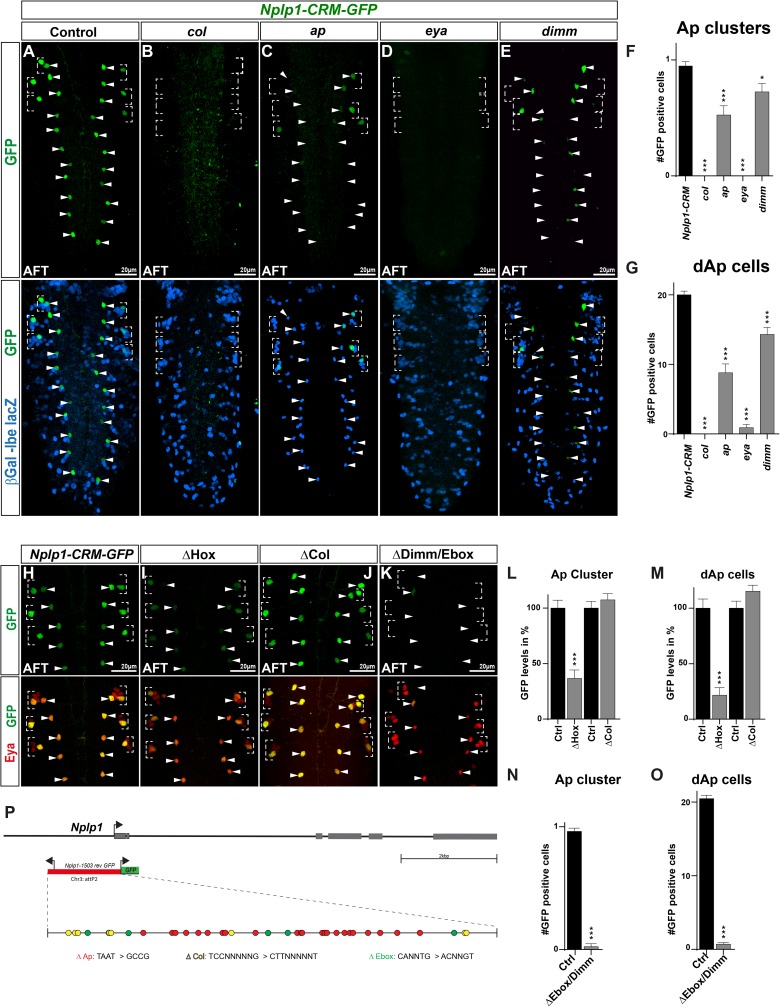
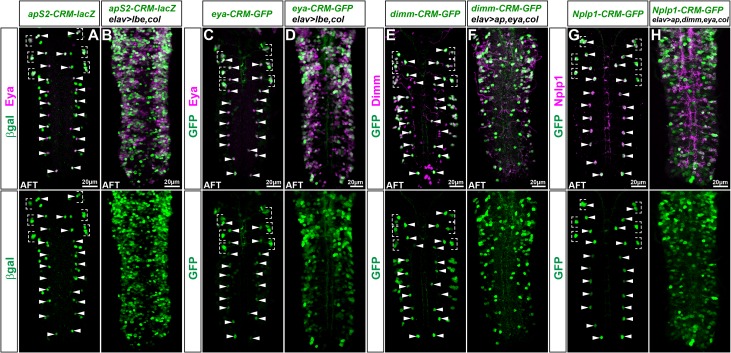
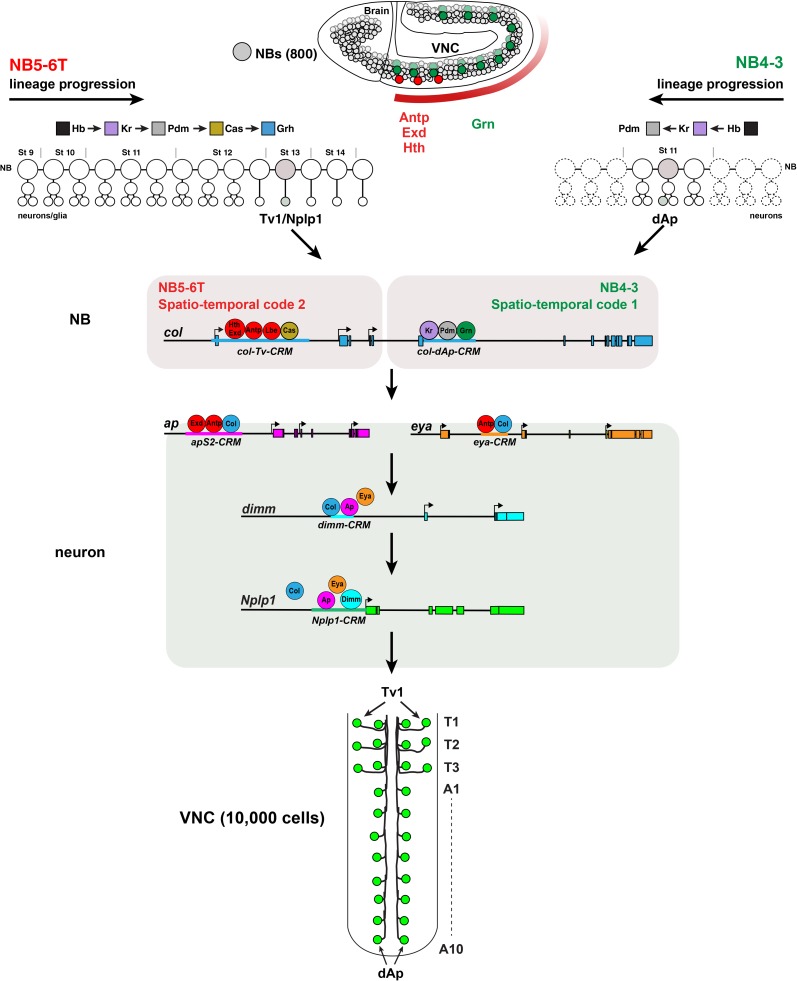
The extensive genetic regulatory flows underlying specification of different neuronal subtypes are not well understood at the molecular level. The Nplp1 neuropeptide neurons in the developing Drosophila nerve cord belong to two sub-classes; Tv1 and dAp neurons, generated by two distinct progenitors. Nplp1 neurons are specified by spatial cues; the Hox homeotic network and GATA factor grn, and temporal cues; the hb -> Kr -> Pdm -> cas -> grh temporal cascade. These spatio-temporal cues combine into two distinct codes; one for Tv1 and one for dAp neurons that activate a common terminal selector feedforward cascade of col -> ap/eya -> dimm -> Nplp1. Here, we molecularly decode the specification of Nplp1 neurons, and find that the cis-regulatory organization of col functions as an integratory node for the different spatio-temporal combinatorial codes. These findings may provide a logical framework for addressing spatio-temporal control of neuronal sub-type specification in other systems.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Hobert O. Regulatory logic of neuronal diversity: terminal selector genes and selector motifs. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(51):20067–71. Epub 2008/12/24. PubMed Central PMCID: PMC2629285. doi: 10.1073/pnas.0806070105 - DOI - PMC - PubMed
-
- Wenick AS, Hobert O. Genomic cis-regulatory architecture and trans-acting regulators of a single interneuron-specific gene battery in C. elegans. Developmental cell. 2004;6(6):757–70. Epub 2004/06/05. doi: 10.1016/j.devcel.2004.05.004 - DOI - PubMed
-
- Sharma K, Sheng HZ, Lettieri K, Li H, Karavanov A, Potter S, et al. LIM homeodomain factors Lhx3 and Lhx4 assign subtype identities for motor neurons. Cell. 1998;95(6):817–28. - PubMed
-
- Thor S, Andersson SG, Tomlinson A, Thomas JB. A LIM-homeodomain combinatorial code for motor-neuron pathway selection. Nature. 1999;397(6714):76–80. doi: 10.1038/16275 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
