

Flexibility and Design: Conformational Heterogeneity along the Evolutionary Trajectory of a Redesigned Ubiquitin
- PMID: 28416112
- PMCID: PMC5415430
- DOI: 10.1016/j.str.2017.03.009
Flexibility and Design: Conformational Heterogeneity along the Evolutionary Trajectory of a Redesigned Ubiquitin
Abstract
Although protein design has been used to introduce new functions, designed variants generally only function as well as natural proteins after rounds of laboratory evolution. One possibility for this pattern is that designed mutants frequently sample nonfunctional conformations. To test this idea, we exploited advances in multiconformer modeling of room-temperature X-ray data collection on redesigned ubiquitin variants selected for increasing binding affinity to the deubiquitinase USP7. Initial core mutations disrupt natural packing and lead to increased flexibility. Additional, experimentally selected mutations quenched conformational heterogeneity through new stabilizing interactions. Stabilizing interactions, such as cation-pi stacking and ordered waters, which are not included in standard protein design energy functions, can create specific interactions that have long-range effects on flexibility across the protein. Our results suggest that increasing flexibility may be a useful strategy to escape local minima during initial directed evolution and protein design steps when creating new functions.
Keywords: directed evolution; high-resolution X-ray crystallography; protein design; room-temperature X-ray crystallography; ubiquitin.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
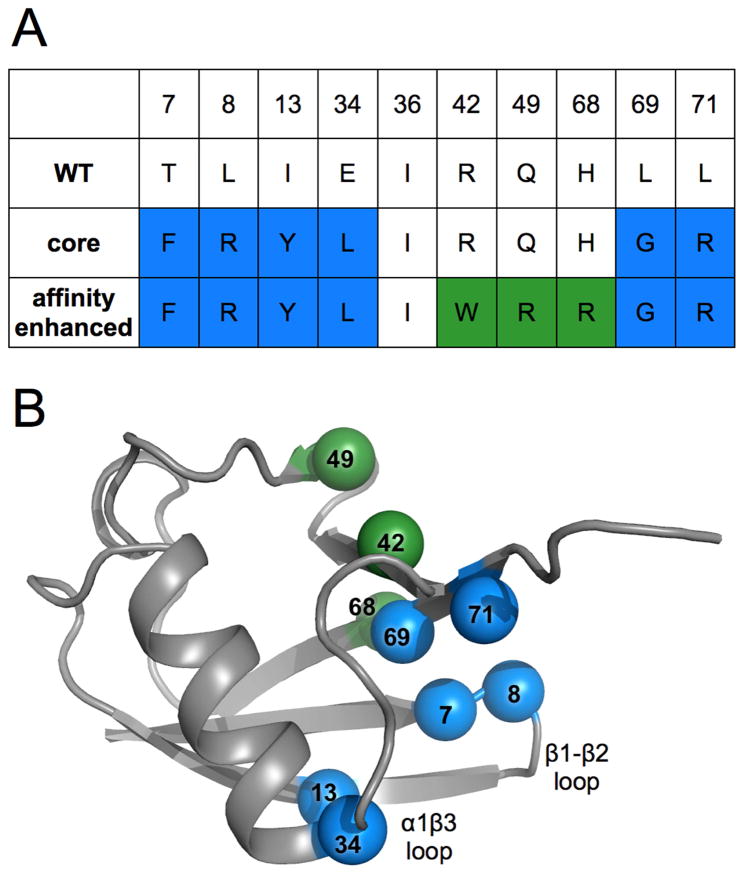
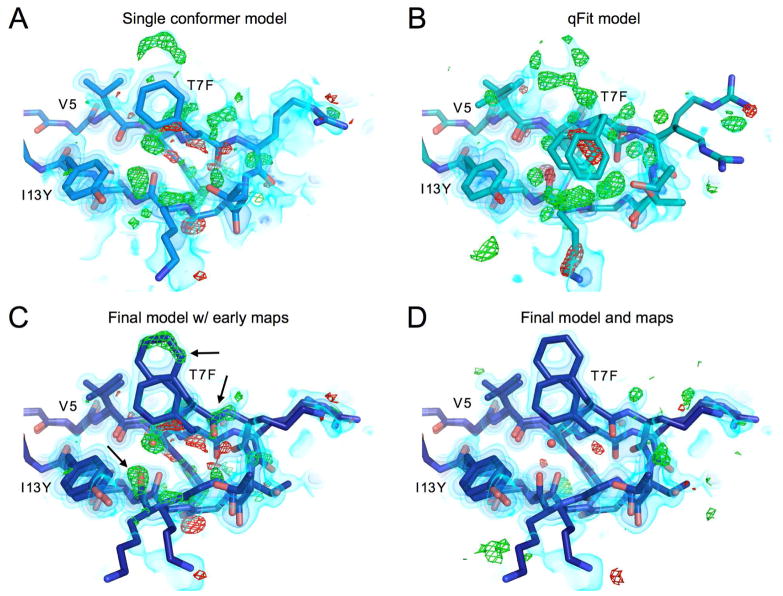
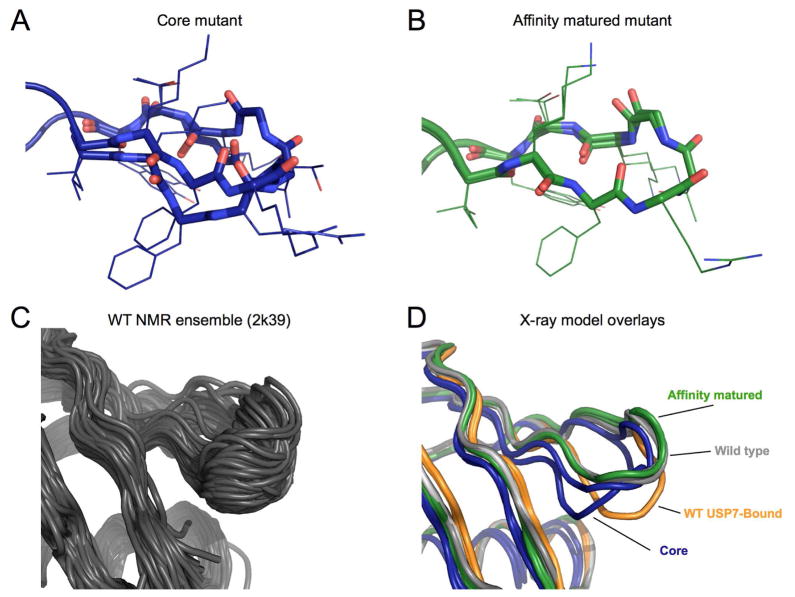
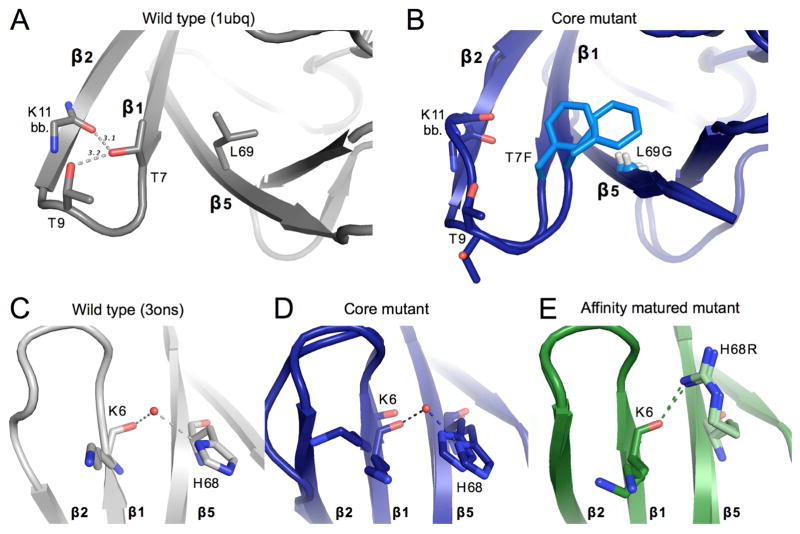
Comment in
-
Room Temperature X-Ray Crystallography Reveals Conformational Heterogeneity of Engineered Proteins.Structure. 2017 May 2;25(5):691-692. doi: 10.1016/j.str.2017.04.004. Structure. 2017. PMID: 28467914
References
-
- Adams PD, Grosse-Kunstleve RW, Hung LW, Ioerger TR, McCoy AJ, Moriarty NW, Read RJ, Sacchettini JC, Sauter NK, Terwilliger TC. PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr D Biol Crystallogr. 2002;58:1948–1954. - PubMed
-
- Blomberg R, Kries H, Pinkas DM, Mittl PRE, Grütter MG, Privett HK, Mayo SL, Hilvert D. Precision is essential for efficient catalysis in an evolved Kemp eliminase. Nature. 2013;503:418–421. - PubMed
-
- Brünger AT, Adams PD, Rice LM. New applications of simulated annealing in X-ray crystallography and solution NMR. Structure. 1997;5:325–336. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
