

Targeting Autocrine CCL5-CCR5 Axis Reprograms Immunosuppressive Myeloid Cells and Reinvigorates Antitumor Immunity
- PMID: 28416485
- PMCID: PMC5484057
- DOI: 10.1158/0008-5472.CAN-16-2913
Targeting Autocrine CCL5-CCR5 Axis Reprograms Immunosuppressive Myeloid Cells and Reinvigorates Antitumor Immunity
Abstract
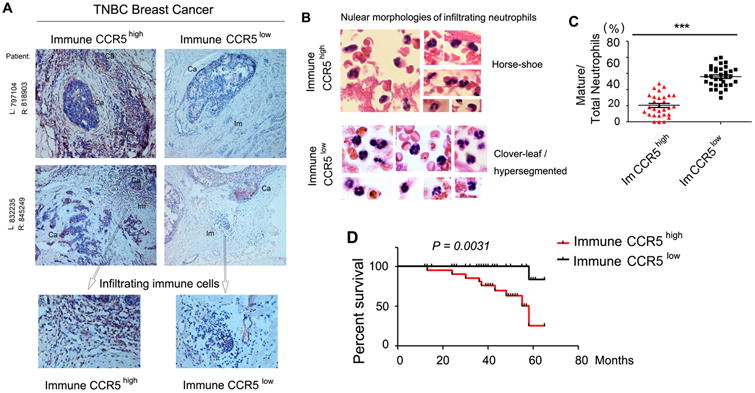
The tumor-promoting potential of CCL5 has been proposed but remains poorly understood. We demonstrate here that an autocrine CCL5-CCR5 axis is a major regulator of immunosuppressive myeloid cells (IMC) of both monocytic and granulocytic lineages. The absence of the autocrine CCL5 abrogated the generation of granulocytic myeloid-derived suppressor cells and tumor-associated macrophages. In parallel, enhanced maturation of intratumoral neutrophils and macrophages occurred in spite of tumor-derived CCL5. The refractory nature of ccl5-null myeloid precursors to tumor-derived CCL5 was attributable to their persistent lack of membrane-bound CCR5. The changes in the ccl5-null myeloid compartment subsequently resulted in increased tumor-infiltrating cytotoxic CD8+ T cells and decreased regulatory T cells in tumor-draining lymph nodes. An analysis of human triple-negative breast cancer specimens demonstrated an inverse correlation between "immune CCR5" levels and the maturation status of tumor-infiltrating neutrophils as well as 5-year-survival rates. Targeting the host CCL5 in bone marrow via nanoparticle-delivered expression silencing, in combination with the CCR5 inhibitor Maraviroc, resulted in strong reductions of IMC and robust antitumor immunities. Our study suggests that the myeloid CCL5-CCR5 axis is an excellent target for cancer immunotherapy. Cancer Res; 77(11); 2857-68. ©2017 AACR.
©2017 American Association for Cancer Research.
Conflict of interest statement
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
