

Coding of navigational affordances in the human visual system
- PMID: 28416669
- PMCID: PMC5422815
- DOI: 10.1073/pnas.1618228114
Coding of navigational affordances in the human visual system
Abstract
A central component of spatial navigation is determining where one can and cannot go in the immediate environment. We used fMRI to test the hypothesis that the human visual system solves this problem by automatically identifying the navigational affordances of the local scene. Multivoxel pattern analyses showed that a scene-selective region of dorsal occipitoparietal cortex, known as the occipital place area, represents pathways for movement in scenes in a manner that is tolerant to variability in other visual features. These effects were found in two experiments: One using tightly controlled artificial environments as stimuli, the other using a diverse set of complex, natural scenes. A reconstruction analysis demonstrated that the population codes of the occipital place area could be used to predict the affordances of novel scenes. Taken together, these results reveal a previously unknown mechanism for perceiving the affordance structure of navigable space.
Keywords: affordances; dorsal stream; navigation; occipital place area; scene-selective visual cortex.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
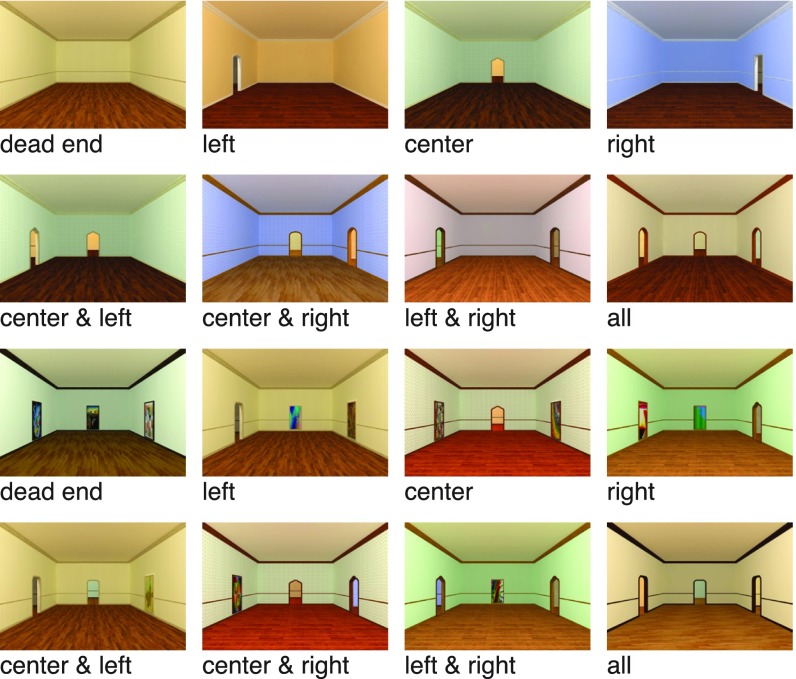

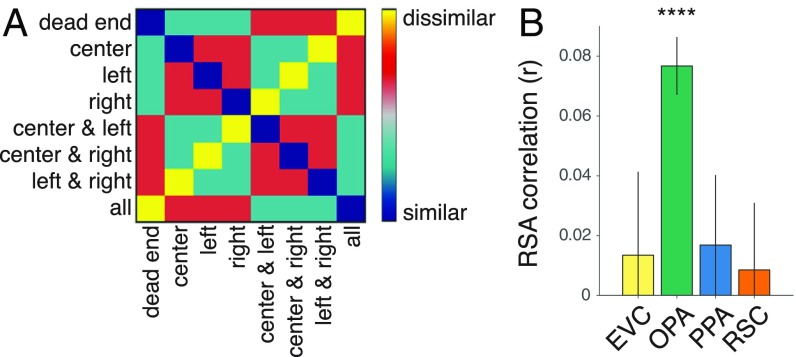
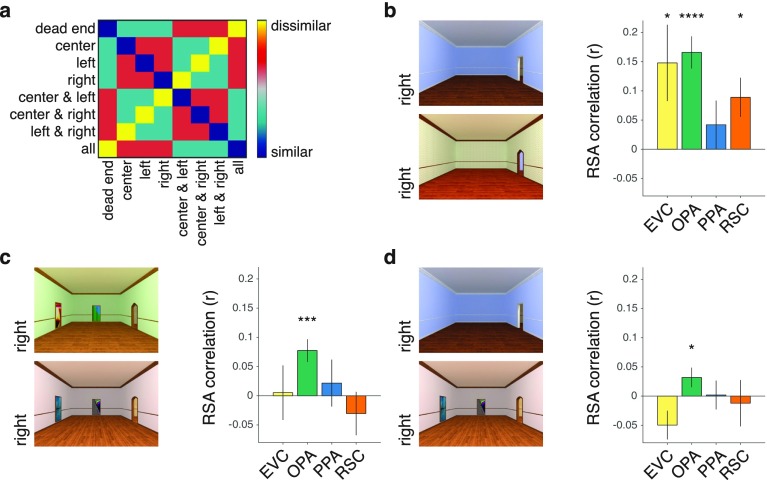
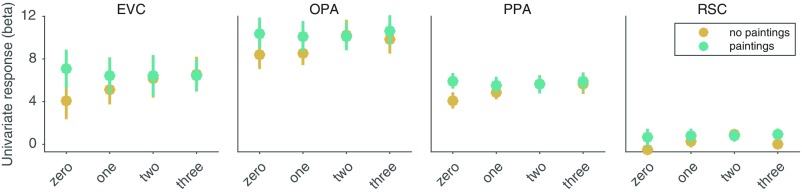



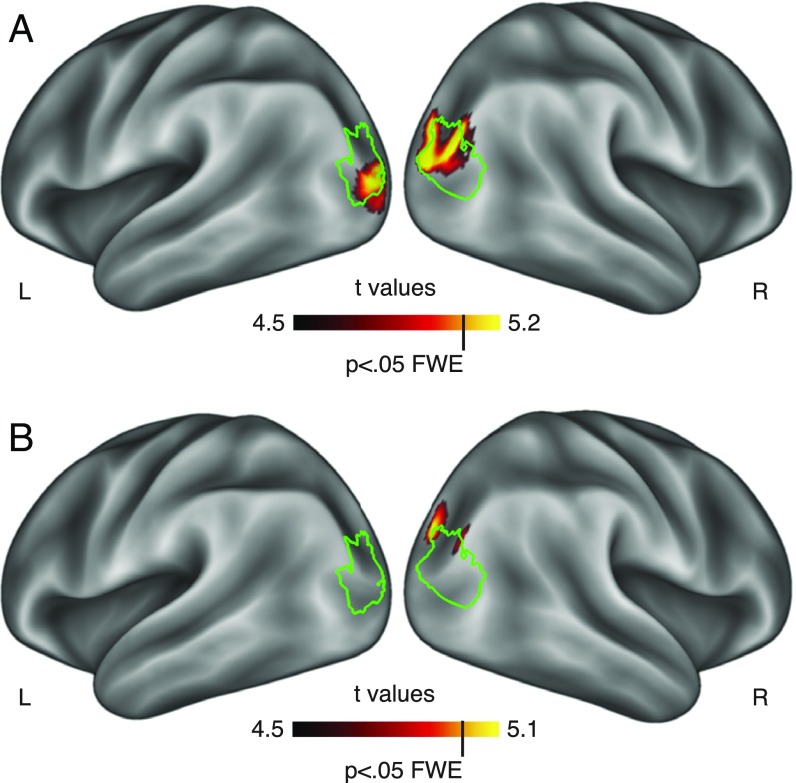
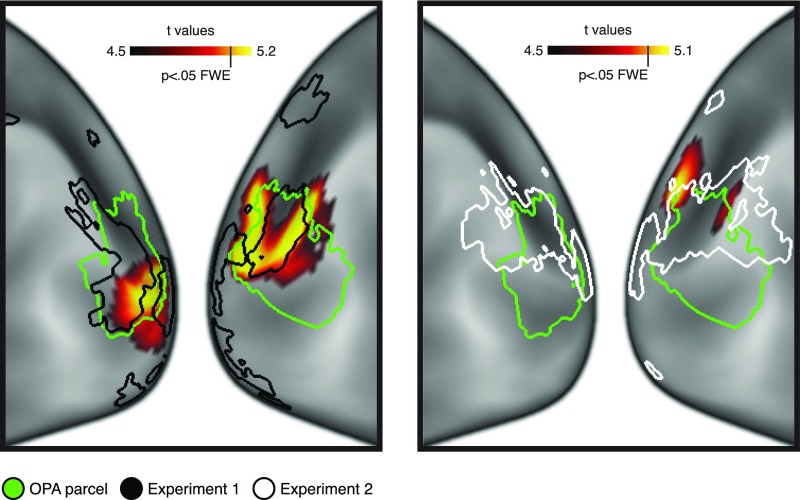
Comment in
-
Human Navigation: Occipital Place Area Detects Potential Paths in a Scene.Curr Biol. 2017 Jun 19;27(12):R599-R600. doi: 10.1016/j.cub.2017.05.012. Curr Biol. 2017. PMID: 28633030
References
-
- Gibson JJ. Visually controlled locomotion and visual orientation in animals. Br J Psychol. 1958;49:182–194. - PubMed
-
- Gibson JJ. The theory of affordances. In: Shaw R, Bransford J, editors. Perceiving, Acting and Knowing. Erlbaum; Hillsdale, NJ: 1977.
-
- Cisek P, Kalaska JF. Neural mechanisms for interacting with a world full of action choices. Annu Rev Neurosci. 2010;33:269–298. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
