

Sensorimotor-independent development of hands and tools selectivity in the visual cortex
- PMID: 28416679
- PMCID: PMC5422761
- DOI: 10.1073/pnas.1620289114
Sensorimotor-independent development of hands and tools selectivity in the visual cortex
Abstract
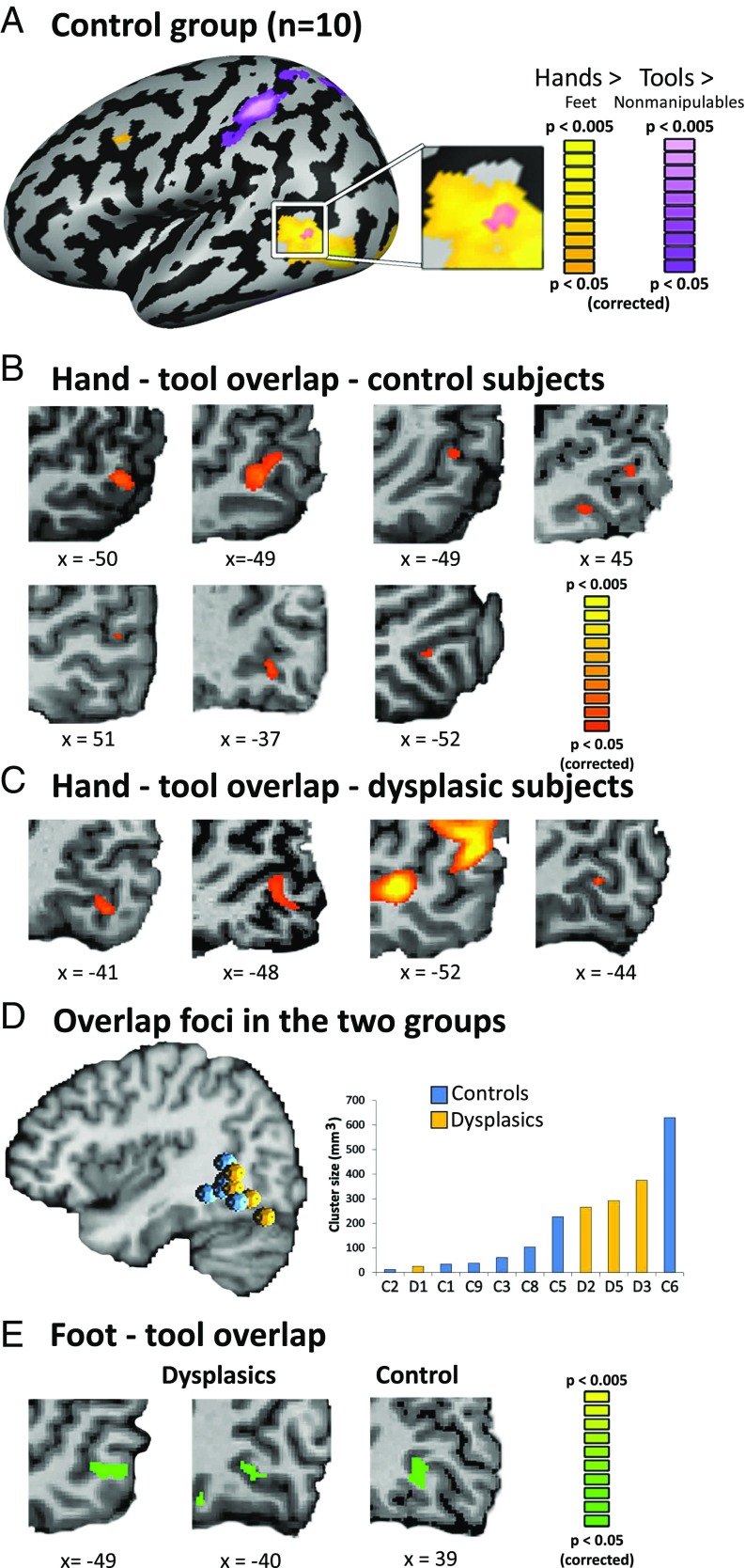
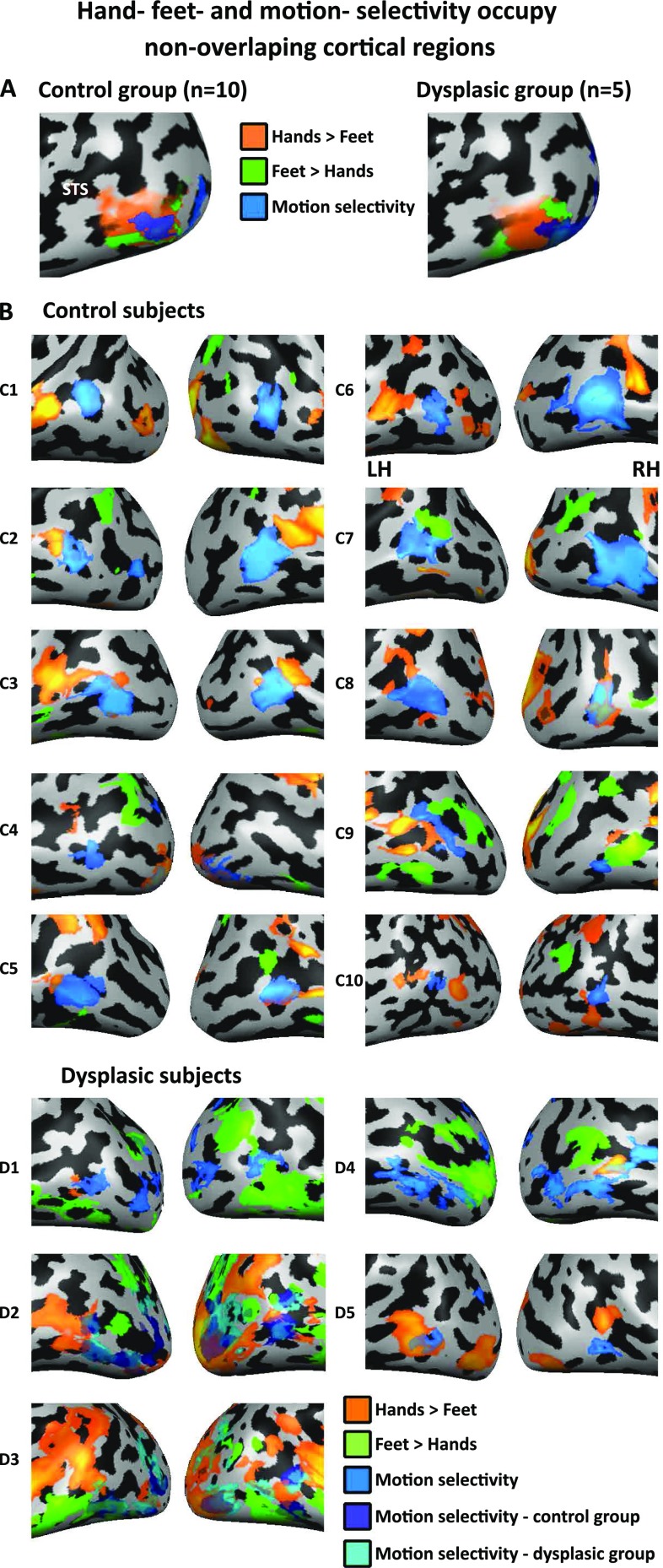
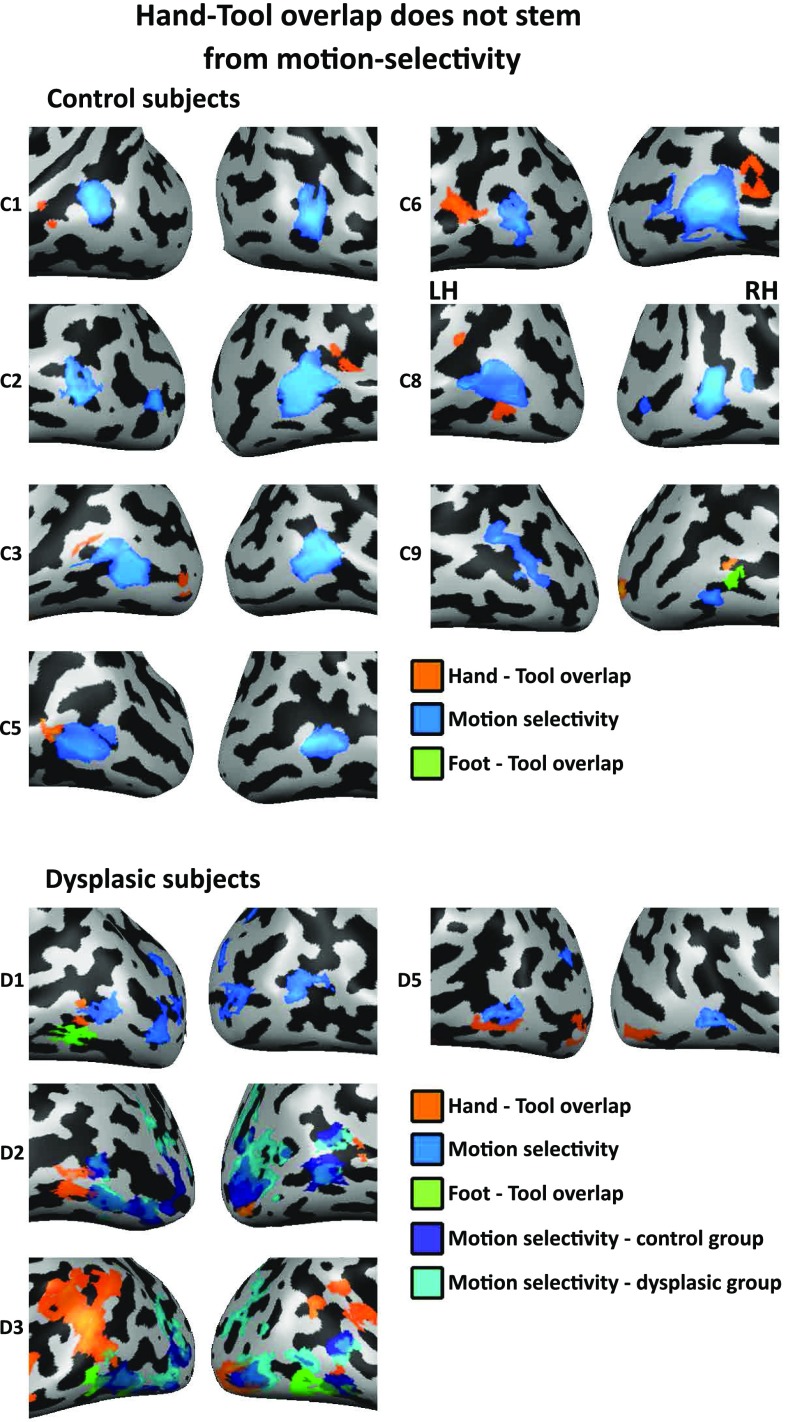
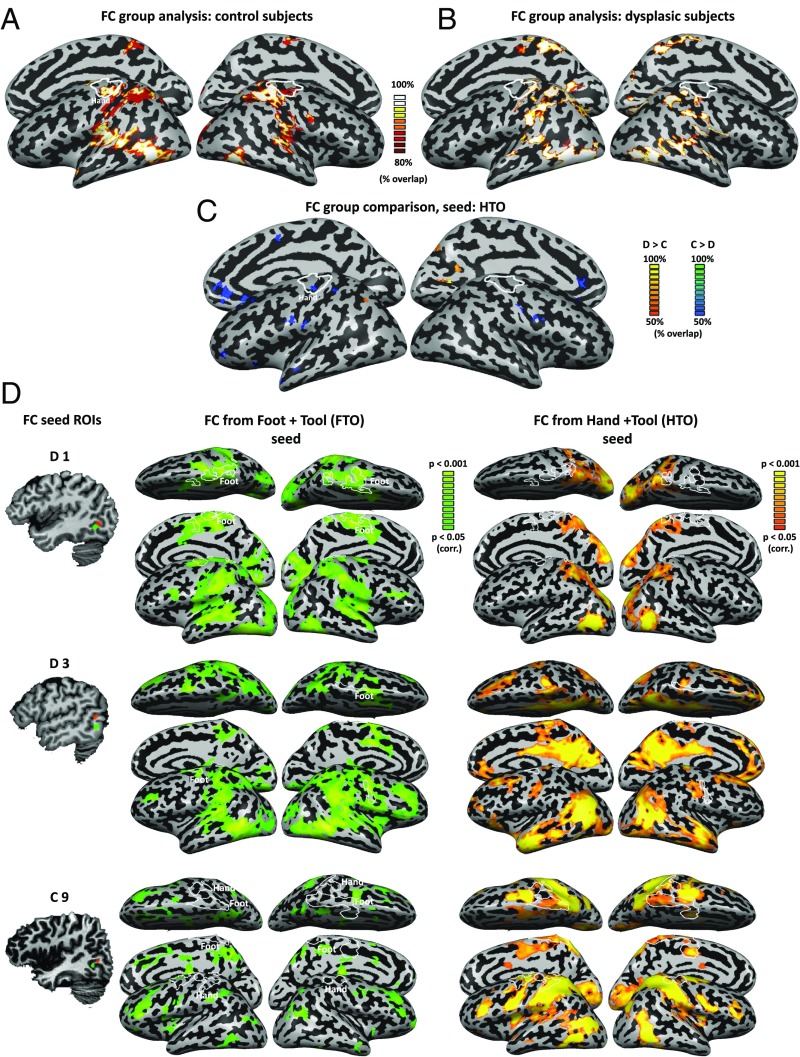
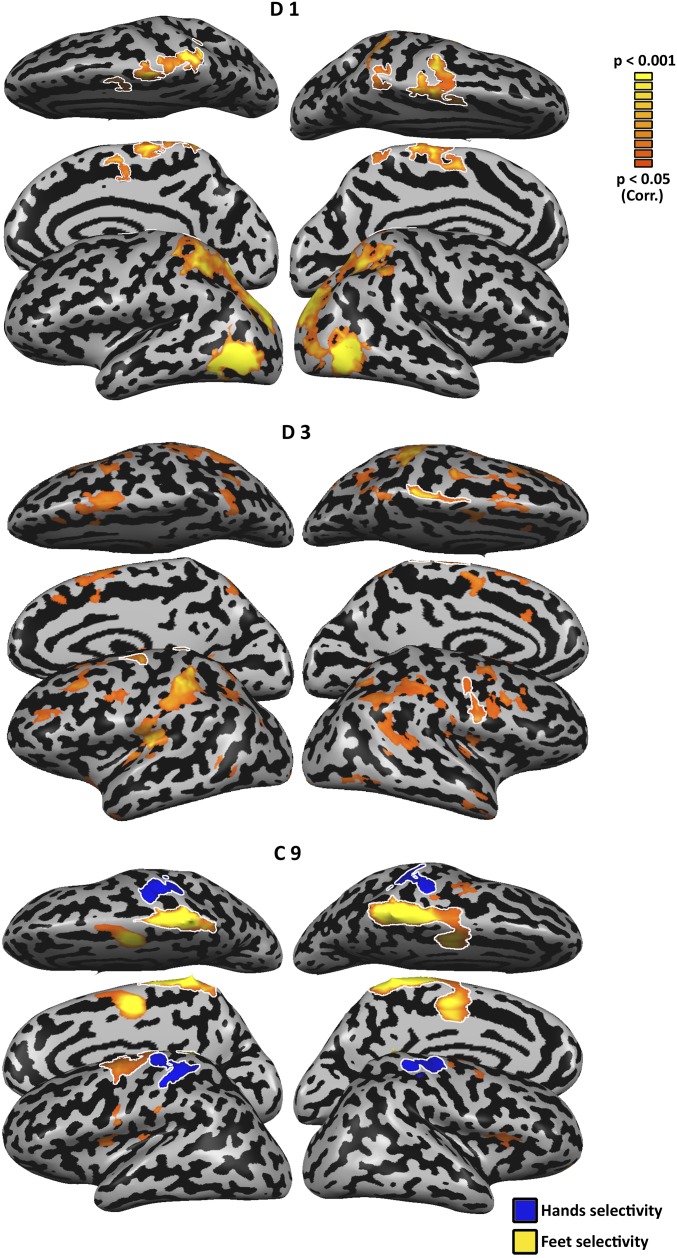
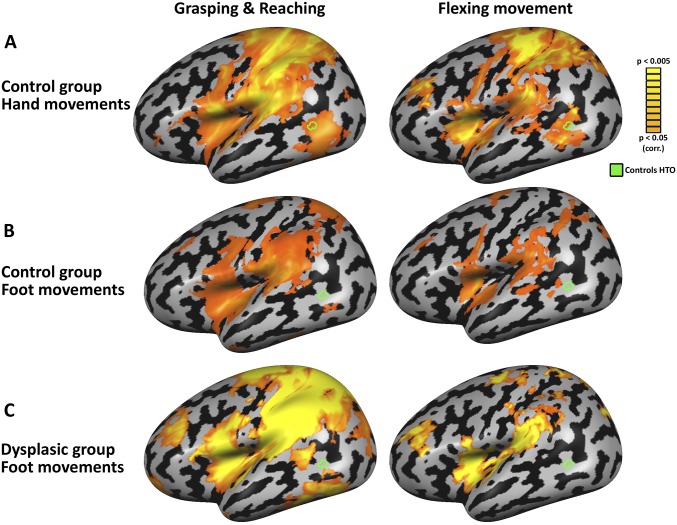
The visual occipito-temporal cortex is composed of several distinct regions specialized in the identification of different object kinds such as tools and bodies. Its organization appears to reflect not only the visual characteristics of the inputs but also the behavior that can be achieved with them. For example, there are spatially overlapping responses for viewing hands and tools, which is likely due to their common role in object-directed actions. How dependent is occipito-temporal cortex organization on object manipulation and motor experience? To investigate this question, we studied five individuals born without hands (individuals with upper limb dysplasia), who use tools with their feet. Using fMRI, we found the typical selective hand-tool overlap (HTO) not only in typically developed control participants but also in four of the five dysplasics. Functional connectivity of the HTO in the dysplasics also showed a largely similar pattern as in the controls. The preservation of functional organization in the dysplasics suggests that occipito-temporal cortex specialization is driven largely by inherited connectivity constraints that do not require sensorimotor experience. These findings complement discoveries of intact functional organization of the occipito-temporal cortex in people born blind, supporting an organization largely independent of any one specific sensory or motor experience.
Keywords: body image; brain development; motor deprivation; tool use; visual cortex.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Plasticity based on compensatory effector use in the association but not primary sensorimotor cortex of people born without hands.Proc Natl Acad Sci U S A. 2018 Jul 24;115(30):7801-7806. doi: 10.1073/pnas.1803926115. Epub 2018 Jul 11. Proc Natl Acad Sci U S A. 2018. PMID: 29997174 Free PMC article.
-
Large-Scale Organization of the Hand Action Observation Network in Individuals Born Without Hands.Cereb Cortex. 2019 Jul 22;29(8):3434-3444. doi: 10.1093/cercor/bhy212. Cereb Cortex. 2019. PMID: 30169751
-
Closely overlapping responses to tools and hands in left lateral occipitotemporal cortex.J Neurophysiol. 2012 Mar;107(5):1443-56. doi: 10.1152/jn.00619.2011. Epub 2011 Nov 30. J Neurophysiol. 2012. PMID: 22131379
-
Human brain plasticity: evidence from sensory deprivation and altered language experience.Prog Brain Res. 2002;138:177-88. doi: 10.1016/S0079-6123(02)38078-6. Prog Brain Res. 2002. PMID: 12432770 Review.
-
Object Domain and Modality in the Ventral Visual Pathway.Trends Cogn Sci. 2016 Apr;20(4):282-290. doi: 10.1016/j.tics.2016.02.002. Epub 2016 Mar 2. Trends Cogn Sci. 2016. PMID: 26944219 Review.
Cited by
-
Novel objects with causal event schemas elicit selective responses in tool- and hand-selective lateral occipitotemporal cortex.Cereb Cortex. 2023 Apr 25;33(9):5557-5573. doi: 10.1093/cercor/bhac442. Cereb Cortex. 2023. PMID: 36469589 Free PMC article.
-
Consensus Paper: Cerebellum and Social Cognition.Cerebellum. 2020 Dec;19(6):833-868. doi: 10.1007/s12311-020-01155-1. Cerebellum. 2020. PMID: 32632709 Free PMC article. Review.
-
Is an artificial limb embodied as a hand? Brain decoding in prosthetic limb users.PLoS Biol. 2020 Jun 8;18(6):e3000729. doi: 10.1371/journal.pbio.3000729. eCollection 2020 Jun. PLoS Biol. 2020. PMID: 32511238 Free PMC article.
-
Towards Strong Inference in Research on Embodiment - Possibilities and Limitations of Causal Paradigms.J Cogn. 2021 Jan 8;4(1):5. doi: 10.5334/joc.139. J Cogn. 2021. PMID: 33506171 Free PMC article.
-
Hand-Selective Visual Regions Represent How to Grasp 3D Tools: Brain Decoding during Real Actions.J Neurosci. 2021 Jun 16;41(24):5263-5273. doi: 10.1523/JNEUROSCI.0083-21.2021. Epub 2021 May 10. J Neurosci. 2021. PMID: 33972399 Free PMC article.
References
-
- Caramazza A, Shelton JR. Domain-specific knowledge systems in the brain: The animate-inanimate distinction. J Cogn Neurosci. 1998;10:1–34. - PubMed
-
- Blundo C, Ricci M, Miller L. Category-specific knowledge deficit for animals in a patient with herpes simplex encephalitis. Cogn Neuropsychol. 2006;23:1248–1268. - PubMed
-
- Proklova D, Kaiser D, Peelen MV. Disentangling representations of object shape and object category in human visual cortex: The animate-inanimate distinction. J Cogn Neurosci. 2016;28:680–692. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
