

A simplified protocol for differentiation of electrophysiologically mature neuronal networks from human induced pluripotent stem cells
- PMID: 28416807
- PMCID: PMC5984104
- DOI: 10.1038/mp.2017.56
A simplified protocol for differentiation of electrophysiologically mature neuronal networks from human induced pluripotent stem cells
Abstract
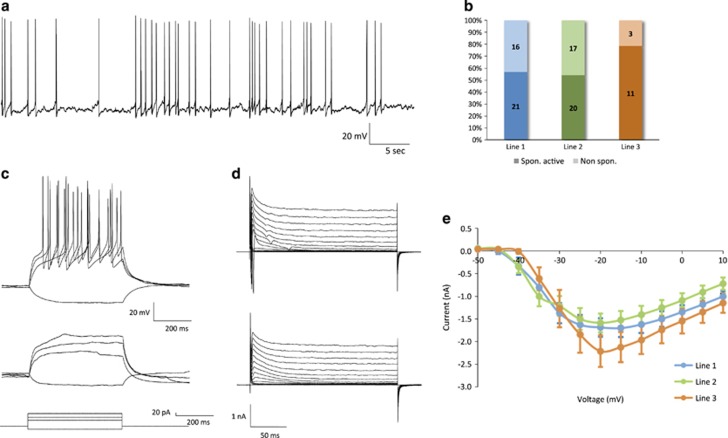
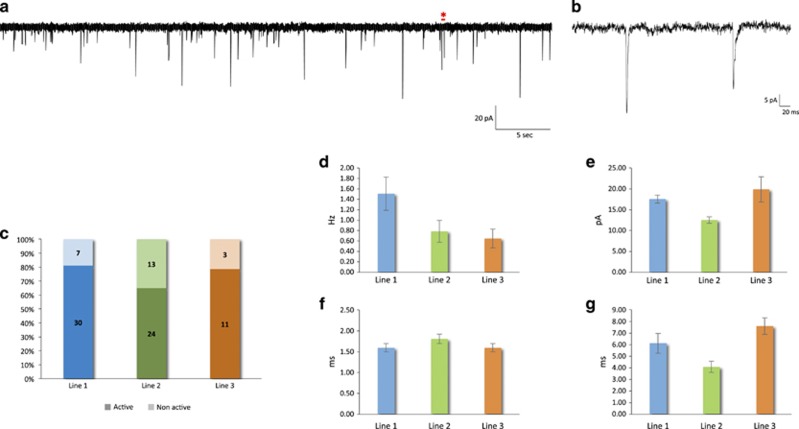
Progress in elucidating the molecular and cellular pathophysiology of neuropsychiatric disorders has been hindered by the limited availability of living human brain tissue. The emergence of induced pluripotent stem cells (iPSCs) has offered a unique alternative strategy using patient-derived functional neuronal networks. However, methods for reliably generating iPSC-derived neurons with mature electrophysiological characteristics have been difficult to develop. Here, we report a simplified differentiation protocol that yields electrophysiologically mature iPSC-derived cortical lineage neuronal networks without the need for astrocyte co-culture or specialized media. This protocol generates a consistent 60:40 ratio of neurons and astrocytes that arise from a common forebrain neural progenitor. Whole-cell patch-clamp recordings of 114 neurons derived from three independent iPSC lines confirmed their electrophysiological maturity, including resting membrane potential (-58.2±1.0 mV), capacitance (49.1±2.9 pF), action potential (AP) threshold (-50.9±0.5 mV) and AP amplitude (66.5±1.3 mV). Nearly 100% of neurons were capable of firing APs, of which 79% had sustained trains of mature APs with minimal accommodation (peak AP frequency: 11.9±0.5 Hz) and 74% exhibited spontaneous synaptic activity (amplitude, 16.03±0.82 pA; frequency, 1.09±0.17 Hz). We expect this protocol to be of broad applicability for implementing iPSC-based neuronal network models of neuropsychiatric disorders.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
